Neuronal Patterning and Regionalization
Neuronal Patterning and Regionalization. Steps during neural development:. Neurogenesis Compartmentalization Neural differentiation Neural migration Axonal guidance Synaptogenesis. Neural development in vertebrate embryo: Gastrulation.


Neuronal Patterning and Regionalization
E N D
Presentation Transcript
Steps during neural development: • Neurogenesis • Compartmentalization • Neural differentiation • Neural migration • Axonal guidance • Synaptogenesis
Neural development in vertebrate embryo: Gastrulation Blastula stage through neurulae, highlighting gastrulationand neurulation. Blastulastage embryo with 3 germ layers, first signs of invagination of dorsal blastopore lip Embryo in midgastrulation, involution of dorsal mesoderm (organizer tissue). Gastrula stage embryo: Embryo at end of gastrulation. The 3 germ layers have arrived at their final destination
Organizing Centers:Restricted specialized areas that are crucial for the induction of area specification • Spemann’s organizer (dorsoblastopore lip) • Hensen’s node (similar to Spemann’s org) • Roofplate and notochord become organizers • secondary organizers: • Isthmic organizer (IsO) • Anterior neural ridge (ANR) • Cortical hem
Organizer Transplant experiment A region just above the blastopore lip (mesodermal tissue) is excised & transplanted to ventral side of host. The host embryo develops a secondary dorsal axis, first evident by a secondary neural plate. A section through a host embryo with two dorsal axes: Secondary dorsal axis contains the same tissues as the primary dorsal axis, including a nervous system. Note: neural tissue was derived from recipient cells, not donor cells. Thus, the transplant had altered the fate of the overlying cells
Default model of neural induction.Balance between agonists and antagonists!Importance of inhibition as a developmental regulatory mechanism. Expression of signaling factors: Bone morphogenic protein (BMP), a TGF-β-like PGF expressed in ectoderm on ventral side, inducing ectoderm to become epidermis. Organizer on the dorsal side releases inhibitors of the BMPs: noggin, chordin, and follistatin, which diffuse into the ectoderm on the dorsal side, block the effects of BMPs, and allow neural tissue to form.
Signaling pathway involving BMPs • Large family of polypeptide growth factors (PGF) related to transforming growth factor-β (TGF-β): BMP, activin, and GDF group members. • Heterodimer receptors, with type I & type II subunits, cytoplasmic domains with serine/theronine kinase activity. • Dimerization after binding of a TGF-β-like PGF starts signal transduction pathway: Activation of cytoplasmic proteins (SMADs), which translocate to nucleus to activate expression of downstream target genes. • Inhibitory mechanisms regulate signaling: • Extracellular proteins such as chordin, tolloid, and twisted gastrulation interact with the BMP-like ligands, regulating their diffusion through the extracellular milieu and their ability to bind receptor • Cell surface proteins such as BAMBI inhibit signaling by binding up BMPs but failing to transduce a signal. • Inhibitory SMADs poison the signal transduction pathway.
Neurulation The neural plate forms after gastrulation is completed. The neural tube narrows along its medial-lateral Axis. The plate begins to role into a tube. The cells at the midline produce a medial hinge point (MHP). As the tube forms and segregates into the embryo, neural crest cells emigrate from the dorsal aspect of the neural tube. G.C. Schoenwolf
Steps during neural development: • Neurogenesis • Compartmentalization • Neural differentiation • Neural migration • Axonal guidance • Synaptogenesis
Pattern Formation In the early stages of pattern formation, two perpendicular axes are established -Anterior/posterior (A/P, head-to-tail) axis -Dorsal/ventral (D/V, back-to-front) axis Polarity refers to the acquisition of axial differences in developing structures Position information leads to changes in gene activity, and thus cells adopt a fate appropriate for their location
AP polarity of vertebrate CNS • Head organizer becomes precordal mesoderm (PME) underneath prechordal plate • Tail organizer becomes notochord and somites, underneath epichordal neural plate
Early Neural Patterning:Establishment of AP Axis • Head and tail organizer release factors which create a gradient.
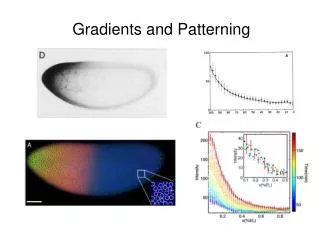
Neural Patterning • A/P polarity and other key organizational features are 1st established by gradients of positional information of a gradient of a substance or signal. • How can a gradient confer positional information? …can signal relative concentrations correlated with distance. A. Evidence of positional info in a single 2-dimensional system; e.g., the striking stripped patterns on insect wings (butterflies, moths). • A chemical signal from eyespot center determine the pigment elaborated by surrounding cells as a function of concentration. • Excision of eyespot center absence of eyespot pigmentation. • Transplantation of an eyespot center to an ectosyne locus stimulates the development of an eyespot in the surrounding tissue.
In the 3-dimensional system of the embryo, the initial establishment of A/P polarity is signalled by the organizer (dorsal lip of the blastopore in amphibians; Hensen’s node in birds). • During gastrulation, the organizer tissues come to underlie the neural plate and differentiate into the notochord. The chordal mesoderm, which underlies the future midbrain, hindbrain, and spinal cord, apparently sends out distance signals from prechordal mesoderm. [These have been reversed in transplantation experiments to demonstrate this].
The candidate neural inducers, which have been studied (chordin, noggin, and follistatin) induce primitive neural tissue that appears to be forebrain-like; chordin particularly potent. Recall that these 3 proteins antagonize members of TGF-β signalling family of molecules. This suggests that induction of anterior neural plate differentiation involves inhibitors of TGF-β-like signals that repress neural development. This would be a “ground state”, which would be induced to be more posterior by a 2nd signal: a transforming signal.
In this case, a type of gradient, a ratio between activating (noggin) and transforming signals would determine the A/P polarity along the neuraxis. Possible candidate “posteriorizers” (transforming signals) include bFGF and retinoic acid.
Tail organizer: FGF, WNT, RA &BMP inhibitors are posteriorizing signaling molecules Head organizer: BMP InhibitorsCordin and Noggin, Wnt inhibitors Cerberus, Dickkopf and frzb1 to "anteriorize" neural tube
Regionalization of the Nervous System • Segmentation (see below) • Developmental control genes (e.g., Hox), which encode positional values along A/P axis. Positional signaling mechanism, which activates these genes may be a (more complex) version of a simple earlier model (gradients?): At Henson’s Node, a strong candidate for this signal is a gradient of retinoic acid, which regulates the pattern of Hox gene expression. Different Hox genes at specific locations respond more or less readily to lower or higher [RA]s, through a family of receptors, which, bound by RA, become transcription factors.
Regarding Segmentation: Subdivision of the main body axis by segmentation is a developmental theme found in many animal phyla. This provides compartments, which allocate precursor cells into a repeated set of similar molecules, so that developmental fields can remain small, and specialization of cell types and patterns can be generated as local variations on the repetitive theme. Mesoderm = segmented into somites, yielding muscle groups. The neuraxis is also segmented ---------------
Rhombomeres – the clearest subdivision partition the hindbrain neuroepithelium. See next slides (Fig. 2.6) for these 8 segments. In the CNS, segmentation is a mechanism for specifying pattern during development. The earliest neurons and neural pathways are laid out in stripes, which match a morphological repeat pattern ( a “2-segment repeat” pattern, which has similar patterns of development in even- or odd-numbered segments). Cf., cerebellum
How do the cells become segregated? • Mechanical boundaries (certain extracellular matrix pattern, such as chondroitin SO4 appear at the boundaries during development (however, only important during later devel.). • Differential adhesion between cells (re-aggregation experiments show that this does indeed occur through a 2-segment repeat rule (evens evens; odds odds), so that adjacent rhombomeres remain separate.
Pattern Generation does not Involve only the Migration of Cells themselves, but also the Axons of Cells • How does a neuronal axon “know” how to travel to a given area and make specific connections? • Appears to involve three steps: • pathway selection • target selection • address selection
Pattern Generation • What role does the substrate play in directing the pathway of axons?
II. Developmental Control Genes. These genes, which encode txn factors, or signaling molecules, are expressed in a spatially variable manner. These classes of genes and their segmented pattern of expression in the rhomomeres are shown in Fig. 2.5 The Hox genes (homeobox family) have a clustered chromosomal organization. [The relative position of the gene reflects the expression along the A/P axis]. This expression of the Hox gene confers positional value and regional identity.
Pattern Generation • Are there other molecules known to direct the migration of axons?
Pattern Generation • What is the function of neurotrophins?
Pattern Generation • What is involved in the formation of a synapse?
Pattern Generation • What is the role of neurotropic factors in the survival of neurons?
Pattern Generation • What is known about the migration of retinal ganglion axons?
Pattern Generation • How do axons distinguish between different regions of optic tectum?
What is the signaling mechanism for expression of these genes? As noted earlier, this is a gradient of RA. The RA signal regulates the pattern of Hox expression. There is a direct correspondence between the location of the Hox gene in its cluster and its responsiveness to RA.
5’ Genes respond less rapidly; require higher [RA]s Genes respond more rapidly at lower [RA]s 3’ Anterior CNS Posterior CNS Change in Hox gene expression change in morphology along the A/P axis
Patterning of the brain and spinal cord through compartmentalization: Melton, Iulianella, Trainor, 2004 Regional patterning: Forebrain (FB), Midbrain (MB), Hindbrain (HB) and Spinal cord (SC). Graded Wnt signaling functions along the entire length of the neuraxis inducing progressively more posterior neural fates. Hox genes play important roles in establishing regional cell identity. This is achieved via opposing gradients of RA and FGF signaling.
Hox gene expression domains in the CNS Nested domains of homeotic genes along the AP axis of the Drosophila and mouse CNS. Hox genes specify a positional value along the AP axis, which is interpreted differently in fly and mouse in terms of downstream gene activation, resulting in neural structure; after Hirth et al., (1998).
Compartmental organization of hindbrain into rhombomeres Stage 8-9: Genes are expressed in alternate stripes that correspond with presumptive Rhombomeres. Stage 9-10: Restriction of movement of mitotic Precursor cells across interfaces. Stage 13: The interfaces between Rhombomeres acquire molecular and Morphological specialization marked by distinct boundaries. Example of odd/even gene expression in Drosophila in situ localization of the achaete transcript From Skeath et al, 1992. Julie E. Cooke, Cecilia B. Moens, 2002
Stages in the compartmental organization of rhombomeres. Genes such as Krox20 and EphA4 (blue) and ephrin-B2 (pink) are expressed in alternate, fuzzy-edged stripes (left). Subsequently, restriction to the movement of mitotic precursor cells occurs at the interfaces between newly formed rhombomeres, which are now sharply defined, and marked by increased intercellular spaces. (right) Sharpening of boundaries and cell lineage restriction occur through the interaction of Eph and ephrin molecules. Data from Fraser et al. (1990).
Regional specification in the developing brain Five-vesicle state Three-vesicle state of a chick embryo
Dorsal Ventral pattern: Notochord as organizer Left: During development, the floor plate (red) develops above the mesodermal notochord (n) and motor neurons (yellow) differentiate in adjacent ventrolateral region of the neural tube. Center: Grafting a donor notochord (n') alongside the folding neural plate results in formation of an additional floor plate and a third column of motor neurons. Right: Removing the notochord from beneath the neural plate results in the permanent absence of both floor plate and motor neurons in the region of the extirpation. Pax6 expression (blue) extends through the ventral region of the cord.
Sonic-hedge-hog expression by notochord & floor plate, control of ventral patterns Shh activity in the ventral neural tube (blue dots) is distributed in a ventral-high, dorsal-low profile within the ventral neural epithelium. 5 classes of neurons are generated in response to graded Shh signalling T.M. Jessell, 2000
Model for ventral neural patterning by SHH. Left: Graded SHH signaling from the ventral pole induces expression of some homeobox genes (e.g., Nkx2.2, Nkx6.1) and represses existing expression of others (e.g. Pax6, Dbx2). Center: Cross-repressive interactions between pairs of transcription factors sharpen mutually exclusive expression domains. Right: Profiles of homeobox gene expression define progenitor zones and control neuronal fate. After Briscoe and Ericson, (2001).
Regulation of DV pattern in the telencephalon by SHH. Cross section of mouse telencephalon at early (left) and later (right) stage. SHH produced in the ventral midline region controls development of basal ganglia primordia and medial and lateral ganglionic eminences (MGE, LGE). First, ventral SHH induces MGE gene expression; SHH (partly produced by the MGE) induces LGE gene expression later.
The neural tube, shown here for a mouse, is subdivided into four longitudinal domains: the floor plate, basal plate, alar plate, and roof plate. Motor neurons are derived from the basal plate.