Enzymes: Kinetics & Inhibition
E N D
Presentation Transcript
Enzyme Active protein acts as a biological catalyst. Since it is a protein, enzyme consists of amino acids. The molecular weight ranges from 15000 to millions Dalton The enzyme reduces the activation energy for the reaction in order to increase the reaction kinetic constant (k). Arrhenius equation:
Enzyme classification • Oxidoreductase: transfer oxygen atoms or electron • Transferase: transfer a group (amine, phosphate, aldehyde, oxo, sulphur, etc) • Hydrolase: hydrolysis • Lyase: transfer non-hydrolytic group from substrate • Isomerase: isomerazion reactions • Ligase: bonds synthesis, using energy from ATPs
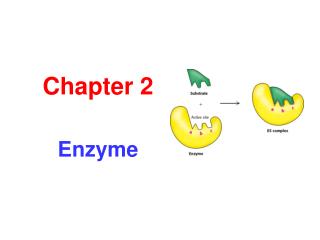
How do enzymes work? Lock and key principle + + Substrate enzyme substrate-enzyme product enzyme complex Many enzymes need cofactor (apoenzymes) such as metal ions or several organic compounds. However, several metal ions and organics might inhibit enzyme kinetics. For example, heavy metals at high dose, EDTA, glucose, etc
The mechanism of enzymatic reactions • The simple schematic of enzymatic reaction: • S + E ES E + P • Assumptions: • The enzyme total concentration is constant • The amount of enzyme <<< substrate • The product concentration is low • (1st order reaction)
Michaelis-Menten kinetic • The approaches: • Rapid equilibrium • Quasy steady state • V: rate of enzymatic reaction • Vm: maximum rate of enzymatic reaction • CS: substrate concentration • Km: substrate affinity constant (Michaelis-Menten constant)
How to obtain the kinetic parameter (Vm, Km)? • Construct and obtain V at different CS. Use graphical methods: • Lineweaver-Burk plot (double reciprocal plot) • Eadie-Hofstee plot • Hanes-Woolf plot • Batch kinetic plot Each plot has its own pros and cons. However, the most popular is double reciprocal plot.
Allosteric enzyme Enzyme with cooperative binding that has more than one active site. Mostly is regulatory enzyme. n: cooperative coefficient
The inhibitions on enzymatic reactions • Inhibitors reduce enzyme activity to substrate. Four type of inhibitions: • Competitive inhibition • Non competitive inhibition • Uncompetitive inhibition • Substrate inhibition • Beside that, the temperature and pH control the enzymatic kinetics. • The pH determines the changes in the ionic form of the enzyme’s active site and changes the enzyme activity. High T may give high rate of reaction, but enzyme thermal denaturation occurs.
Immobilized enzymes • The restriction of enzyme mobility in a fixed space • Advantages: • Enzyme reutilization • Elimination of enzyme recovery and purification • May provide better environment for enzyme activity • Disadvantages: • Problem in mass transfer • Enzyme leakage into solution • Reduced enzyme activity and stability • Lack of controls on micro environmental conditions
Batch culture • Batch growth consists of: • Lag phase • Acceleration phase • Exponential growth phase • Deceleration phase • Stationary phase • Death phase
Lag phase The lag phase occurs immediately after innoculation and is a period of adaption of cells to a new environment. Microorganisms reorganize their molecular constituents when they are transferred to a new medium. The internal machinery of cells is adapted to the new environmental conditions. During this phase, cells mass may increase a little, without an increase in cell number density. Pseudo-lag phase may occur if the inoculum is small or poor inoculum conditions. The long lag phase is possible if the nutrient and growth factors concentrations are low. Additionally, the lag period increases with the age of inoculum. The multiple lag phase may be observed when the medium contains more than one carbon source diauxic growth
Log or exponential phase In this phase, the cells have adjusted to the new environment. Cell mass and cell number density increase exponentially with time. This is the period of balance growth in which all components of cells grow at the same rate. The exponential growth rate is first order , CX: cell concentration at t = t , : specific growth rate CX0: cell concentration at t = 0 At this phase, mostly = max.
Stationary phase • This phase starts at the end of deceleration phase, when the net growth rate is zero or when the growth rate is equal to the death rate. The cells are still metabolically active and produce secondary metabolites (nongrowth-related). During this phase: • Total cell mass concentration may stay constant, but the number of viable cells may decrease • Cell lysis may occur, viable cell mass may drop causing cryptic growth (growth on lysis products of lysed cells) • Secondary metabolite (some hormones, antibiotics) may be produced as a result of metabolite deregulation
Stationary phase The endogenous metabolism occurs where cells catabolizes cellular reserves for new building blocks and for energy-producing monomers. The cells must always expend energy to maintain an energized membrane (pmf), transport of nutrients, and for essential metabolic functions such as motility and repairing. This energy expenditure is called maintenance energy. The reason for termination of growth may be either exhaustion of an essential nutrient or accumulation of toxic products.
Death phase The death phase follows the stationary phase. However, some cell death may start during stationary phase, and a clear demarcation between these two phases is not always possible. The rate of death usually follows first-order kinetics: , NS: concentration of cells at the end of stationary phase N: concentration of cells at t time after stationary phase k’d: 1st order death rate constant
Monod kinetics • Assumptions: • Only single chemical species is growth rate limiting • No inhibitions either by substrates or products • The maintenance coefficient is negligible : specific growth rate, max: maximum specific growth rate KS: saturation constant, CS: substrate concentration
Monod kinetics The Monod kinetic is similar to Michaelis-Menten kinetic The double reciprocal plot (1/ vs. 1/CS) is also used to determine KS and max. However, usually max is obtained by plot CX vs.t.
Other kinetics • Several more complicated kinetics are available, e.q: • Blackman equation • Tessier equation • Moser equation • Contois equation • Multiple substrate equations • Model with growth inhibitors • Logistic equation, etc
Rates & yields Specific rate (qi): ri: rate of production/ consumption of component i Thus, Yield is defined as the ration of production of compound i to the consumption of the compound j (Yji). Some literatures use Yi/j. grams of cells produced /grams of substrate consumed
Maintenance coefficient Cells consume substrate for growth, production, and maintenance. Some part of substrate is used for maintenance such as maintain an energized membrane (pmf), transport of nutrients, and for essential metabolic functions such as motility and repairing. The maintenance coefficient is independent to .
Continuous culture in chemostat The continuous culture provide constant environment to cells in order to obtain prolonged periods of growth and product formation. At certain time, the system usually reaches steady state where cells, products, and substrates concentrations are constant.
The mass balances in chemostat The cells mass balance: D: dilution rate (hour-1) The substrate mass balance: Usually = 1
The mass balances in chemostat The cells mass balance explains that the specific growth rate is controlled by defining the feed flow rate. Integrating with Monod equation: CS: residual substrate concentration The biomass concentration in gram dry weight/liter: Cso: inlet substrate concentration Cxo = 0
Critical dilution rate The dilution rate has the critical value, Dcrit max. It is slightly lower than max. If D > Dcrit, cell is washed out, CX = 0. Then, the substrate is not consumed, CS = Cso.
Comments on batch process • , YSX, qS are constant at max, YSXmax, qSmax (during log phase) • Microbial model parameters are easily determined by combining the exponential equation with CS and CX data vs. time. • , the most important rate, can not be controlled in batch culture
Comments on continuous process • All rates are defined. D = is defined, thus –qS, YSX, qP, qO2, etc are set. • Residual substrate concentration is independent to CSo. • A good experimental tool to study and apply: • Kinetics, to obtain YSXmax, mS, qSmax, KS. • Physiological of microorganisms under defined steady state by changing substrate or electron acceptor or type of medium limitation • Waste water treatment • Industrial fermentation (not widely applied) • Genetic and metabolic studies
Growth monitoring methods • Can be direct or indirect • Petroff-Hauser counting chamber. Rapid but cannot distinguish live and dead cells. Needs a high cell concentration • Electronic cell counters. Use electrical resistance principle. Errors can occur due to cell clumping and the presence of particulate debris • Plate counting technique. Counting cell colony, which is difficult • Turbidity and spectrophotometric techniques. Measure the light scattered or absorbed by the cells. Most popular method, rapid, but not good for colored culture • Dry weight estimation. Uses vacuum filtration and oven to obtain dry cells. Other suspended non-cellular material could give error • On line estimation. Measuring O2, CO2 to estimate biomass concentration • ATP bioluminometry. Measure cell concentration by measuring ATP because ATP is rapidly lost from dead cells.