Receptors and transduction 1
350 likes | 640 Vues
Receptors and transduction 1. Dr. MV Hejmadi. References: Chapter 11 – Neuron by Levitan & Kaczmarek OR Chapter 6 – Neuroscience by Purves et al K. Tsuzuki and T. Ozawa. Glutamate receptors www.els.net

Receptors and transduction 1
E N D
Presentation Transcript
Receptors and transduction 1 Dr. MV Hejmadi References: Chapter 11 – Neuron byLevitan & KaczmarekOR Chapter 6 – Neuroscience byPurves et al K. Tsuzuki and T. Ozawa. Glutamate receptorswww.els.net Madden, D. Structure and function of glutamate receptor ion channels (2002). Nature Revs Neurosci. 3, 91.
Synapse: the point where electrical conduction is converted to chemical conduction (mostly)
The Chemical Synapse 5) Ligand binding 6) Depolarization 7) transmitter recycling 8) vesicular membrane recycling 1) AP invades 2) Ca++ influx 3) Depolarization release of NT 4) NT diffusion
Excitatory synaptic transmission by L-glutamate Action potential Calcium channels Calcium ions L-glutamate vesicles L-glutamate release AMPAR Na+ NMDAR Mg2+ Presynaptic Terminal Synaptic cleft Postsynaptic spine
Glutamate Acetylcholine etc GABA
Specificity of responses Following activation by a NT, how does a neuron know what the response should be? Depending on the type of NT (excitatory or inhibitory), key receptors transduce this signal and dictate the nature of the response
Glu ACh DA Substance P GABA Output Each neuron uses one NT to send a signal (usually) • i.e. glutamatergic, GABAergic, cholinergic etc Synaptic inputs to a GABA-ergic medium spiny neurone
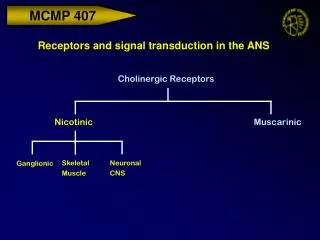
Each NT can activate diverse receptors e.g. acetylcholine can activate 2 classes of receptors, nicotinic or muscarinic Nicotinic AChR • Mimicked by nicotine (agonist) • Only in certain tissues (skeletal muscle, parts of the CNS and PNS) Muscarinic AChR • Mimicked by muscarine (agonist) • Cardiac muscle, cholinergic synapses of the CNS
Nicotinic AChR blocked by antagonists a-bungarotoxin curare Muscarinic AChR Blocked by antagonists like atropine How do you measure specificity? Pharmacological tools Bungarus multicinctus Henbane plant
Receptors categorised based on their transduction mechanisms • Ionotropic receptorsIonotropic receptors are directly coupled to ion channels and are activated by neurotransmitters which open the channel pore to allow movement of specific ions, creating postsynaptic potentials FAST DIRECT B) Metabotropic receptorsThese receptors are not directly coupled to their ion channels and transduce the signal via guanyl nucleotide-binding proteins (G-proteins) that activate intracellular second messenger pathways SLOW INDIRECT PSPs
FAST DIRECT PSPs SLOW INDIRECT PSPs Fig 5.22; Purves et al
Ionotropic receptors Fig 11.8; Levitan and Kaczmarek
Ionotropic receptor types glutamate tetrameric pentameric Assembly of either similar subunits (homomeric) or different subunits (heteromeric)
Heterogeneity of nAChRin mammalian tissues Brain addiction analgesia anxiety attention cognition 10 Autonomic Ganglia cardiovascular gastro-intestinal Muscle contraction
Ionotropic receptors - Generic structure Example:Nicotinic acetylcholine receptor (pentameric) Each subunit has four hydrophobic regions (TMI – IV) Subunits
Ionotropic receptors-Generic structure Unlike other iR, iGluR have a re-entrant loop inTMII domain extracellular N-terminus and intracellular C-terminus Long loop between TMIII and IV forms part of the binding domain with the C-terminal half of the N-terminus glutamate
The modular nature of iGluR subunits Extracellular NTD followed by S1 - sequence between NTD and TMI S2 - sequence between M3 and M4 TMIII and TMIV domains with an intervening re-entrant P loop,. C terminus intracellular S1 and S2 half-domains form the ligand-binding domain. The structure of this domain is shown as a ribbon diagram. It consists of two lobes (lobe I, blue; lobe II, red), separated by a ligand-binding cleft. Lobe 1- structural unit attached to the amino-terminal domain Lobe 2 - structural unit that links to M1 and M3 Madden, D. Structure and function of glutamate receptor ion channels (2002). Nature Revs Neurosci. 3, 91.
Modular structure of iGluRs transmembrane topology, the two extracellular domains, and the carboxyl-terminal domain. Flip/flop refers to an alternatively spliced segment that plays a role in desensitization. S1 refers to the sequence between the amino-terminal domain and M1; S2 refers to the sequence between M3 and M4. Lobe 1 is the structural unit attached to the amino-terminal domain, and Lobe 2 is the structural unit that links to M1 and M3. Note that segments of S1 and S2 are found in both Lobes 1 and 2. Emerging structural explanations of ionotropic glutamate receptor function ROBERT L. McFEETERS and ROBERT E. OSWALD The FASEB Journal. 2004;18:428-438
The 'dimer-of-dimers' model of iGluR assembly Monomers associate most strongly through interactions between their amino-terminal domains (NTDs) (star in middle figure). Dimers undergo a secondary dimerization, mediated by interactions in the S2 and/or transmembrane domains (stars in right-hand figure). The crystallographically observed S1S2 dimer probably corresponds to this secondary dimerization interaction.
NMDA receptor (N-methyl- D-apartate) AMPA receptor(a-amino 3hydroxy 4methyl 5propionic acid) Kainate receptors(kainic acid) Modulates LTP Results in calcium entry, changes in secondary messages altered protein synthesis Epileptogenesis Synaptic plasticity? Flip/flop region-spliced variants (exon 14 –flop and Exon 15 for flip)
The NMDA receptor is blocked by a Mg++ ion at resting potential Calcium as a signal Glutamate directly gates NMDA and AMPAR. NMDAR controls movement of Ca 2+ Na+ and K+ binding sites for glycine, zinc, phencyclidine (angel dust) and Mg2+. Under resting conditions Mg2+blocks the site and keeps the receptors silent. Upon depolarisation the Mg is released and there is movement of Ca into the cell.
NR2 NR1 GLYCINE GLUTAMATE NR2 ? NR1 2+ + Ca Na POLYAMINE 2+ Zn NR1 NR2 OUT MK801 2+ Mg IN NR1-1a/b-4a/b NR2A calmodulin NR2B P P CamKII NR2C NR2D PSD proteins NR3A P = multiple sites for phosphorylation NMDA receptors
So what are the most important NT in the mammalian brain? • glutamate and GABA are the most abundant which mediate synaptic transmission in the CNS via ionotropic receptors (LGICs) • In general, • GABA - inhibitory whereas • glutamate - excitatory
Roles of glutamate in the CNS • Synaptic transmission (EPSPs) • Long term depression • Long term potentiation • Dendritic sproutin • Synaptic modification • Control of gene expression Excess of glutamate release can cause neuronal death (excitotoxicity) e.g. during stroke, epilepsy, Parkinson’s disease
Effects of glutamate-induced excitotoxicity Nature Reviews Neuroscience4; 399-414 (2003
GABA receptors GABA - major inhibitory NT in the mammalian CNS GABA receptors are pentameric in structure 3 classes GABAA and GABAC receptors are ionotropic, GABABreceptors are metabotropic
GABAA receptor binding sites Benzodiazepine – allosteric agonist - tranquilisers / anticonvulsants Barbiturates – prolong action of GABA - anaesthetics / hypnotics
So why have multiple iR? • The post-synaptic response to stimulation can be modulated in the short term (i.e. for hundreds of milliseconds) or for the long term (hours, days or even weeks!!). • Synaptic strength is increased or decreased by altering the level of post-synaptic depolarisation. This is achieved through changing how well receptors respond to stimulation, • by altering the length of time they are active, • the number of receptors physically present or • by altering the amount of L-glutamate that is released into the synaptic cleft