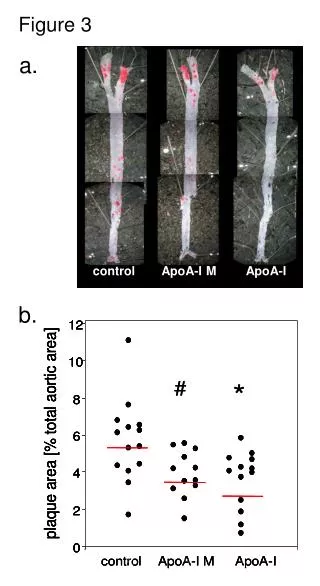
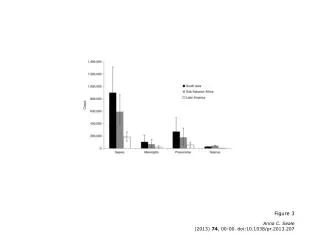
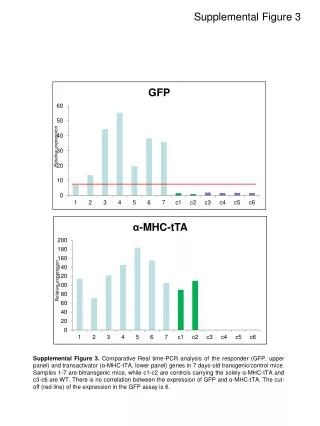
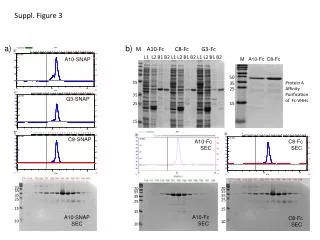
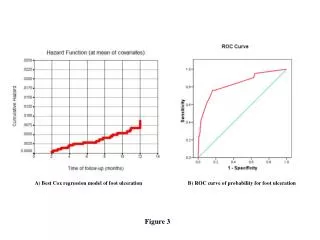
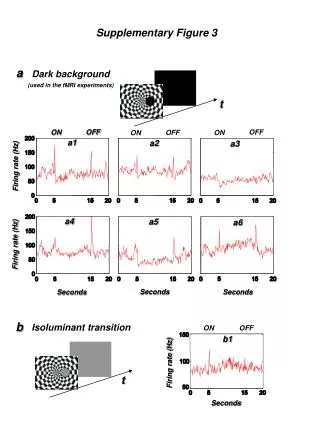
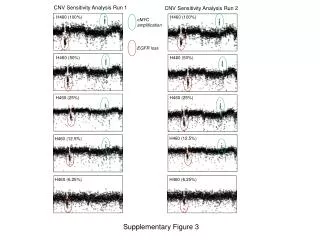
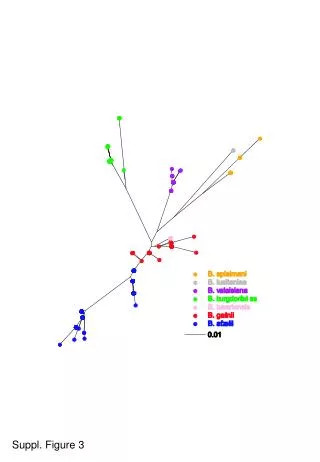
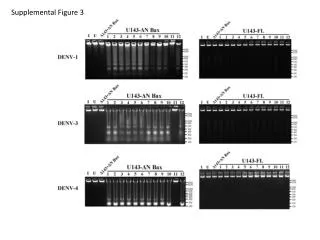
Figure 3
Figure 3 . Purpose: To further characterize cell-to-cell transmission of α -synuclein using an in vitro coculture model Figure 3(A) Hypothesis:


Figure 3
E N D
Presentation Transcript
Figure 3 • Purpose: To further characterize cell-to-cell transmission of α-synuclein using an in vitro coculturemodel • Figure 3(A) Hypothesis: If myc-tagged α-synuclein from donor cells can be released and transmitted to SH-SY5Y acceptor cells, thenα-synuclein will be detected in the donor cells via immunofluorescence.
Figure 3 SH-SY5Y SH-SY5Y SH-SY5Y + α-synuclein myc Q myc myc myc Q Q myc Q Q Acceptor Cells Donor Cells
Figure 3(A): Results • After 24 hrs, myc-tagged α-synuclein from donor cells was detected in acceptor cells • Formation of inclusion bodies in some acceptors cells
Figure 3(A): Results • After 24 hrs, myc-tagged α-synuclein from donor cells was detected in acceptor cells • Formation of inclusion bodies in some acceptors cells *, p<0.05 **, p< 0.01 Conclusions?
Figure 3(B): Results • Inclusion body formation occurs with prolonged transmission of α-synuclein Conclusions?
Figure 3(C) • ~ ½ of the acceptor cells displayed ubiquitin immunoreactivity Figure 3(D) • Cell-to-cell transmission occurs without cellular membrane leakage Conclusions?
Supplemental Fig. S3A • Purpose: To determine if transmission of α-synucleinaggregates is dependent on endocytosis • Dynamin-1 K44A expressed in acceptor cells (blocks endocytic formation) • Donor cells cocultured with acceptor cells • Hypothesis: If transmission of α-synucleinaggregates is dependent on endocytosis, then we would detect a reduction in the uptake of α-synucleinin the cells expressing dynamin-1 K44A.
Fig. S3A: Results • Transmission of α-synuclein significantly reduced in acceptor cells **, p< 0.01 Conclusions?
Figure 4 • Increased α-synuclein accumulation by lysosomal failure but no effect on proteosomal inhibition Conclusions?
Figure 5 • Purpose: To examine the toxicity of endocytosed neuron-derived extracellular α-synuclein to the neurons • Hypothesis: If secreted/endocytosed α-synuclein is toxic to the cells, then we would expect to detect neuronal degeneration and caspase 3 activation.
Figure 5 (A & B): Results • Neurons showed nuclear fragmentation and increased immunoreactivity of activated caspase 3 over 3 days *, p<0.05 **, p< 0.01
Figure 5 (C): Results • MCNSCs showed activated caspase 3 immunoreactivity within 4 weeks *, p<0.05 Conclusions?
Overall Conclusions 1. There is evidence for direct cell-to-cell transmission of α-synuclein. 2. Nerve cells that overexpress tagged α-synuclein can transmit the protein to neural stem cells both in vivo and in vitro. 3. Inclusion body formation occurs with prolonged transmission of α-synuclein. 4. α-synuclein accumulation is achieved when protein quality control systems of the acceptor cells are inhibited. 5. Acceptor neuronal stem cells exhibit cell death and activated caspase 3 due to α-synuclein propagation.
So, why do we care? • Is Parkinson’s Disease really a problem of protein aggregation? • PD is characterized by neuron death and the surviving neurons show aggregations of α-synuclein • If α-synuclein aggregates into an insoluble Lewy Body, PD develops
So, why do we care? • Deposition of Lewy-like inclusions from host to grafted neurons in human PD patients is age-dependent • Understanding how to control α-synuclein aggregation in & around neurons could lead to revolutionary treatment of Parkinson’s Disease • How can we treat PD (based on Desplats et al.’s paper?)