Download

1 / 54
610 likes | 1.02k Vues
Chapter 5 Organization and Expression of Ig Genes. Oct 26 & 31, 2006. 你需要學習的課題 : 抗體基因是如何組成的? 抗體基因重組 (rearrangement) 的機制 抗體的多樣性 (diversity) 是如何產生的? 細胞膜上的抗體如何轉變為分泌性抗體? 抗體的類別 (class) 如何變換? - class switching. Central Feature of Ab Molecules: Vast diversity of Ab specificities

E N D
Chapter 5 Organization and Expression of Ig Genes Oct 26 & 31, 2006
你需要學習的課題: • 抗體基因是如何組成的? • 抗體基因重組 (rearrangement) 的機制 • 抗體的多樣性 (diversity) 是如何產生的? • 細胞膜上的抗體如何轉變為分泌性抗體? • 抗體的類別 (class) 如何變換? - class switching
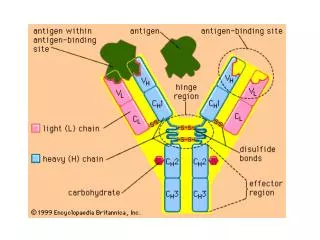
Central Feature of Ab Molecules: • Vast diversity of Ab specificities • A variable (V) region at the N-terminal end and a constant (C) region at the C-terminal end of Ab molecules • Different classes (or isotypes) of Ab (e.g., IgG and IgM) with identical V-region sequences (antigenic specificity)
The Two-gene model of Dryer and Bennett (1965) Two separate genes encode a single Ig H or L chain, one gene for the V region and the other for the C region.
The suggestion that two genes encoded a single polypeptide contradicted the existingone gene-one polypeptideprinciple and was without precedent (先例) in any known biological system.
Verification of the Dryer and Bennet Hypothesis (by Tonegawa and Hozumi, 1976) First direct evidence that separate genes encode the V and C regions of Ig and that the genes are rearranged in the course of B-cell differentiation. - Tonegawa was awarded the Nobel Prize for this work in 1987.
Demonstration of DNA Deletion at an Ig Locus Non-B cells: sperm or liver cells B-cells
Demonstration of DNA Deletion at an Ig Locus deleted sequence 比 大,因此在電泳 時跑得比較慢
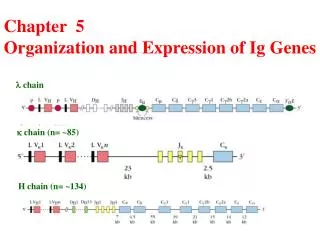
l-Chain Multigene Family Mouse: V region: 2 Vl gene segments 4 Jlgene segments (3 are functional) C region: 4 Clgene segments Human: 30 Vl, 4 Jl and 4 Cl segments
κ-Chain Multigene Family Mouse: V region: ~ 85 Vk gene segments 5 Jkgene segments (4 are functional) C region: 1 Ckgene segment Human: 40 Vk, 5 Jk and 1 Ck segments
H-Chain Multigene Family Mouse: V region: ~ 134 VH gene segments 13 DH gene segments 4 JHgene segments C region: 8 CHgene segments Human: 51 VH, 27 DH, 6 JH and 9 CH segments
V-Region Gene Rearrangements • - The H-chain V-region genes rearrange first, • then the L-chain V-region genes. • The rearrangements occur in an ordered sequence, • but they are random events. • The arrangements of Ig and TCR genes are the • only known site-specific DNA rearrangements • in vertebrates.
H-Chain DNA Undergoes V-D-J Rearrangements (1st rearrangement) (2nd rearrangement) A mature , immunocompetent B cell expresses both IgM & IgD withidentical antigenic specificityon its surface.
L-Chain DNA Undergoes V-J Rearrangements introns are removed
Mechanism of V-region DNA Rearrangements
Two unique recombination signal sequences (RSSs) flanking each germ-line V, D, and J gene segment One-turn RSS: located at 3’ to each Vk, 5’ to each Jl, and both sides of each DHgene segment Two-turn RSS:located at 3’ to each Vl & VH and 5’ to each Jk & JHgene segment
Vλ Jλ CACAGTG 23 nt ACAAAAACC GTGTCAC 12 nt TGTTTTTGG //
One turn/two-turn joining rule The rule ensures that VH, DH, and JH segments join in proper order and that segments of the same type do not join each other.
Gene Segments Are Joined by Recombinases • - Recombination-Activating Genes: RAG-1, RAG-2 • - The proteins encoded by RAG-1 and RAG-2 act • synergistically and are required to mediate • V-(D)-J joining. • Terminal deoxynucleotidyl transferase (TdT), • another lymphoid-specific gene product, is also • involved in V-(D)-J rearrangement.
Process of Recombination of Ig Gene Segments Terminal deoxy-nucleotidyl Transferase Double Strand Break Repair
Defects in Ig-Gene Rearrangements RAG-1-/- or RAG-2-/- mice: - lack RAG-1 or RAG-2 - cannot start the recombination process SCID (severe combined immunodeficiency) mice: - lack double strand break repair (DSBR) enzymes - can carry out synapsis, introduce d.s. breaks - cannot properly join the coding sequences
Ig-gene Rearrangements May Be Nonproductive • Imprecise Joining • - productive and nonproductive • rearrangements • productive rearrangement in • one allele is enough • If rearrangement is not • produced, the B cell dies by • apoptosis. !! !!
Only 1/3 attempts at VL – JL joining, and 1/3 subsequent attempts at VH – DHJH joining, are productive. As a result, < 1/9 (11%) of the early-stage pre-B cells in the bone marrow progress to maturity and leave the bone marrow as mature immunocompetent B cells.
Allelic Exclusion Ensures a Single Antigenic Specificity A single B cell is only specific for a single epitope !!! Once a productive rearrangement is attained, its encoded protein is expressed and the presence of this protein acts as a signal to prevent further gene rearrangement. (1) (2) * active alleles
Antibody Diversity Seven means of generation of Ab diversity: 1. Multiple germ-line V, D, and J gene segments 2. Combinatorial V-(D)-J joining 3. Junctional flexibility 4. P-region nucleotide addition (P-addition) 5. N-region nucleotide addition (N-addition) 6. Somatic hypermutation 7. Combinatorial association of light and heavy chains
Junctional Flexibility Adds Diversity - 4 different joinings of Vk21- Jk1 in pre-B cell lines (Flexible) (Precise)
SinceCDR3makes a major contribution to Ag binding by the Ab molecule, amino acid changes generated by junctional flexibility can make a major contribution to Ab diversity.
P-Addition Adds Diversity at Palindromic Sequences {Palindromic sequences}
N-Addition Adds Considerable Diversity by Addition of Nucleotides add new (N) -nucleotides
- Up to 15 N-nucleotides can be added to both • the DH - JH and VH - DHJH joints. • Thus, a complete H - chain V region is encoded • by a VHNDHNJH unit. • N regions appears to consist of wholly random • sequences
P-nucleotide 及N-nucleotide addition 有些什麼優缺點?
Somatic Hypermutation Adds Diversity in Already-rearranged Gene Segment - Somatic hypermutation occurs only within germinal centers, structures that form in secondary lymphoid organs within a week or so of immunization with an Ag that activates a T-cell-dependent B-cell response. - Somatic hypermutation occurs at a frequency approaching 10-3/bp/generation. This rate is at least 100,000-fold higher than the spontaneous mutation rate, about 10-8/bp /generation, in other genes. - B cells with higher-affinity Ig receptors will be preferentially selected for survival because of their greater ability to bind to the Ag. ----- Affinity Maturation
Experimental Evidence for Somatic Mutation in V region of Ig Genes Most of the mutations are clustered in theCDR1andCDR2hypervariable region.
Antibody Diversity Seven means of generation of Ab diversity: 1. Multiple germ-line V, D, and J gene segments 2. Combinatorial V-(D)-J joining 3. Junctional flexibility 4. P-region nucleotide addition (P-addition) 5. N-region nucleotide addition (N-addition) 6. Somatic hypermutation – after Ag stimulation 7. Combinatorial association of light and heavy chains
Class Switching Among C-Region Genes
Organization of H chain V region C region After antigenic stimulation of a B cell, the H-chain DNA can undergo a further rearrangement in which the VHDHJH unit can combine with any CH gene segment. This process is called class switching.
Class (isotype) switching • Class-specific switch recombinases may • bind to switch regions and facilitate DNA • recombination. • - Cytokines secreted by activated TH cells • have been shown to induce B cells to class • switch to a particular isotype. • - IL-4, for example, induces class switching • from Cm to Cg1 and then from Cg1 to Ce.
Switch regions Class Switching from Cm to Cg1 Class Switching from Cg1 to Ce a circular excision product
Co-expression of membrane forms of m and d H-chains by Alternative RNA Processing
Expression of Membrane or Secreted Ig mRNAs 先暫時不考慮Cd的表現 (sIgM) (mIgM)
Therefore, processing of an Ig H-chain primary transcript can yield different mRNAs, which explains how a single B cell can produce secreted or membrane-bound forms of a particular Ig and simultaneously express IgM and IgD.