The Cytoskeleton
The Cytoskeleton. Tim Mitchison timothy_mitchison@hms.harvard.edu. Cytoskeleton lectures. Intoduction – diverse protein polymers physically organize cells and promote motility Polymerization dynamics Self-assembly only: intermediate filaments

The Cytoskeleton
E N D
Presentation Transcript
The Cytoskeleton Tim Mitchison timothy_mitchison@hms.harvard.edu
Cytoskeleton lectures • Intoduction – diverse protein polymers physically organize cells and promote motility • Polymerization dynamics • Self-assembly only: intermediate filaments • Self-assembly + NTP hydrolysis: Actin and microtubules • Nucleation: controlling where and when polymers form • Motor proteins: mechanical work from ATP hydrolysis • Muscles • Cilia and flagella • Cell division
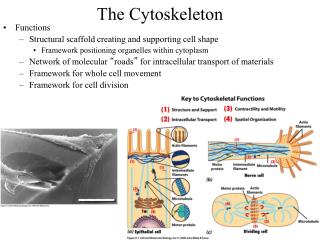
Why should a PhD student be interested in the cytoskeleton? • The cytoskeleton challenges us to consider problems of spatial and temporal organization at scales of m and seconds, where molecular dynamics turn into life. • The cytoskeleton plays fundamental roles in biological processes • Spatial organization of the cell, including organelles and signaling pathways. • Force generation for movement inside cells, and tissue morphogenesis • The cytoskeleton is an often under-appreciated component of the physiology of specialized cell types, and the pathology associated with many diseases. • Cytoskeleton questions often drive development of novel microscopy and single-molecule biophysics technology (and vice versa!) • Direct medical applications • Cytoskeleton poteins can be druggable disease targets • Synthetic cell biology will require spatial organization of cells
Frog egg dividing Each cycle takes ~20min 1 mm
Whole cell length scales Xenopus laevis early embryo Ostreococcus tauri Hendersen et al 2007 E coli ~10-6 m ~10-6 m ~10-3 m
Cleaving eggs solve simple geometry problems 1) centering 2) orientation q ~1200mm How is the 1st cleavage plane positioned so as to accurately bisect the egg? How is the 2nd cleavage plane oriented orthogonal to the 1st?
About how big is a typical human cell? • ~1 mm • ~10 mm • ~100 mm
About how big is a typical protein molecule? • ~1 nm • ~10 nm • ~100 nm • ~1 mm
About how long does it take for a protein sized molecule to diffuse across a ~10mm cell? • ~ 0.01 sec • ~ 0.1 sec • ~ 10 sec • ~ 1000 sec
How can protein-sized molecules organize micron-sized cells?
Organizing cells using molecules Long protein polymers Mechanical forces on membranes Chemical gradients Chemical or electrochemical waves These mechanisms scale in different ways with cell size
Protein polymers physically organize cells and promote motility For the polymer to effectively integrate over space, its ends must have special properties relative to its middle. Structural polarity might also be very useful The biochemistry of the polymer must be compatible with cellular time, length and force scales
Actin filaments; actin Plasma membrane deformation, contraction, migration
Microtubules; tubulin - - - - Mitosis, organelle transport + + + +
Intermediate filaments; keratin, vimentin, neurofilaments, othersAlso nuclear lamins Mechanical integrity. Nuclear organization (nuclear lamins) Role in specifying tissue-specific cell function??
Prokaryote cytoskeleton • Until recently it was thought that bacteria did not have, or need, a cytoskeleton • Small cell size, rigid cell wall no need for internal organization or physical strength • This view has been changed by progress on FtsZ (tubulin relative), MreB & ParM (actin relatives), Crescentin (IF-like), etc (many more) • So far, bacteria have polymers, but not motor proteins • Bacteria cytoskeleton polymers are involved in cell shape, cell division and DNA segregation – like eukaryotes. But, it is far from clear exactly how they function, and whether eukaryotic analogies hold Bacterial actins and their diversity. Ozyamak E, Kollman JM, Komeili A. Biochemistry. 2013 Oct 8;52(40):6928-39. Multidimensional view of the bacterial cytoskeleton. Celler K, Koning RI, Koster AJ, van Wezel GP. J Bacteriol. 2013 Apr;195(8):1627-36. Bacterial cytokinesis: From Z ring to divisome. Lutkenhaus J, Pichoff S, Du S.Cytoskeleton (Hoboken). 2012 Oct;69(10):778-90. Evolution of cytomotive filaments: the cytoskeleton from prokaryotes to eukaryotes. Löwe J, Amos LA. Int J Biochem Cell Biol. 2009 Feb;41(2):323-9.
FtsZ (prokaryotic tubulin relative) Cell division in prokaryotes & chloroplasts, but not mitochondria Note: function in cell division very different from microtubules!
MreB: an actin-like protein involved in cell wall biosynthesis MreB-GFP in B Subtilis Confocal imaging Garner et al Science 333:222
MreB: an actin-like protein involved in cell wall biosynthesis MreB-GFP in B Subtilis Confocal imaging Garner et al Science 333:222 Cell wall Cell wall biosynthesis enzymes Plasma membrane Coupling proteins Precursor transport proteins MreB filaments
How is the MreB moving? 1) Unidirectional (Coherent) 2) Bidirectional (Incoherent) Why does this matter?
Model for MreB function in organizing cell wall biosynthesis MreB oligomers
Bacterial cytoskeleton • Many outstanding fundamental questions • Excellent research groups at HMS and FAS • Druggable targets for antibiotics?
Nematode sperm cells Nematode sperm are ameboid, not flagelated
Major Sperm Protein (MSP) • MSP is a small protein, unique to nematode sperm cells • - MSP polymerization-depolymerization promotes cell crawling • MSP is actin-like in function, but not in structure • no ATP binding, non-polar polymer • Stewart and Roberts (2005) Adv Protein Chem. 71:383-99.
Why is MSP interesting? MSP polymerization-depolymerization drives amoeboid motility of nematode sperm, that resembles amoeboid motility driven by actin polymerization-depolymerization in many eukaroytes (eg Neutrophils, Dictyostelium) Similarities in dynamic organization are striking: polymerization at the leading edge, backwards treadmilling of a cross-linked network that can couple to the substrate, depolymerization near the nucleus Yet, aspects of actin we consider central to this biology (polar filaments, polymerization coupled to ATP hydrolysis) are not present in MSP This challenges us to consider what aspects of cytoskeleton strucutre and dynamics are truly fundamental to cell migration and cell polarity. It also challenges us to think about how a cytoskeleton that can drive cell motility and phagocytosis might evolve
Basic Principles In Cytoskeleton Biology • Self-assembly • Polarity • Polymer assembly: nucleation vs. elongation • Polymer dynamics, critical concentration • Role of NTP hydrolysis: treadmilling and dynamic instability • Filament binding proteins and their diverse functions • Motor proteins: converting chemical energy into mechanical work
Self-assembly Spontaneous assembly of subunits (typically proteins) to build an ordered structure The size and shape of the final structure are governed by the shape and interactions of the subunits Examples of self-assembled structures: Cytoskeleton filaments, virus coats, bacterial flagella, protein machines. Also, pathogenic protein aggregates (Hemoglobin-S polymers, Prions, Alzheimer’s plaques)
The main driving force for protein self-assembly is: • Van der Waals interactions • Electrostatic interactions • Hydrogen bonds • Hydrophobic interactions • All the above
Polarity: the property of having two different ends. • Filament polarity. • Polar filament. Each asymmetric subunit points the same way. The two ends are different and the filament lattice has inherent directionality
Polarity: the property of having two different ends. • Filament polarity. • Polar filament. Each asymmetric subunit points the same way. The two ends are different and the filament lattice has inherent directionality • Non-polar filament. Polypeptides are always asymmetric, but they can polymerize into filament where the two ends are the same and the lattice has no inherent directionalty
Polarity: the property of having two different ends. • Filament polarity. • Polar filament. Each asymmetric subunit points the same way. The two ends are different and the filament lattice has inherent directionality • Non-polar filament. Polypeptides are always asymmetric, but they can polymerize into filament where the two ends are the same and the lattice has no inherent directionalty • Cell Polarity Dendrites Axon Retracting tail Leading edge Basal Apical Migrating fibroblast Neuron Epithelial cells
Filament polarity: cytoskeleton polymers • Polar • Actin filaments • (Some) Prokaryotic actin-related filaments (MreB, ParM) • Microtubules • FtsZ (prokaryotic tubulin-related polymer) • Non-polar • Intermediate filaments, nuclear lamins • Major Sperm protein filaments in nematode sperm • “Bipolar” • Myosin II filaments (muscle, non-muscle) • Eg5 tetramer (Kinesin involved in mitosis) Myosin II minifilament Sinard et al (1989) JCB109:1537
ATP ADP + Pi Filament polarity: significance? • Proteins that bind to the side of the filament will point he same way • Only polar filaments can act as directional tracks for motor proteins
ATP ADP + Pi Filament polarity: significance? • Proteins that bind to the side of the filament will point he same way • Only polar filaments can act as directional tracks for motor proteins • Different protein surfaces are exposed at the two ends • End-specific nucleation • End-specific capping + nucleating factor
Measuring polarity 1) Use proteins that stick to the side in one orientation (EM) Pointed end Barbed end Actin filament Myosin heads An analogous method for microtubules is called “hook decoration”
Measuring polarity 1) Use proteins that stick to the side in one orientation (EM) Pointed end Barbed end Actin filament Myosin heads An analogous method for microtubules is called “hook decoration” 2) Use differential polymerization rate Plus end Minus end Rhodamine-tubulin, GTP
Measuring polarity 1) Use proteins that stick to the side in one orientation (EM) Pointed end Barbed end Actin filament Myosin heads An analogous method for microtubules is called “hook decoration” 2) Use differential polymerization rate Plus end Minus end Rhodamine-tubulin, GTP 3) Use motor protein Minus end Plus end Kinesin + ATP moves towards plus end
Measuring polarity 4) Image a microtubule tip-tracking protein GDP-tubulin GTP-tubulin - +
Measuring polarity GFP 4) Image a microtubule tip-tracking protein in living cell EB1 GDP-tubulin GTP-tubulin - + The tip-tracking protein EB1 binds preferentially to GTP-tubulin in microtubules - +
EB1 imaging in Xenopus egg extract 60x TIRF Ani Nguyen
+ + + + + + + Filament polarity : microtubules in cells Microtubule Motile cell eg fibroblast - + end (Preferred end for subunit addition) - end
+ + + + + + + - - - - + + + + Filament polarity : microtubules in cells Microtubule Motile cell eg fibroblast - + end (Preferred end for subunit addition) - end Epithelial cell Axon - + - + - + Note: situation more complex in dendrites. Polarity depends on distance from soma, and can be mixed
Neuron - + - + - + Filament polarity : vesicle transport + Microtubules direct vesicle trafficking by acting as tracks for motor proteins. Golgi and lysosomes move towards minus ends while secretory vesicles move towards at plus ends Motile cell eg fibroblast + + + + + + - - - - Epithelial cell + + + +
Filament polarity : actin filaments in cells Barbed end (Preferred end for subunit addition) pointed end In the leading edge of a migrating cell, actin filaments are organized with their barbed ends pointing forwards. As the membrane protrudes, new actin subunits polymerize onto these barbed ends Filopodium: long, thin rod with bundled actin Lamellipodium: thin sheet with dendritic actin Migrating fibroblast