The Cell Cycle
Molecular Biology II. The Cell Cycle. The Life Cycle of Cells. The Cell Cycle Follows a Regular Timing Mechanism. Newly born cells grow and perform their functions. After reaching a specific point in their life cells divide via mitosis to give rise to daughter cells.


The Cell Cycle
E N D
Presentation Transcript
Molecular Biology II The Cell Cycle
The Life Cycle of Cells • The Cell Cycle Follows a Regular Timing Mechanism. • Newly born cells grow and perform their functions. • After reaching a specific point in their life cells divide via mitosis to give rise to daughter cells. • Cell Differentiation Creates New Types of Cells. • Cells Die by Suicide
The size difference between a neuron (from the retina) and a lymphocyte in a mammal Both cells contain the same amount of DNA. A neuron grows progressively larger after it has permanently withdrawn from the cell cycle. During this time, the ratio of cytoplasm to DNA increases enormously (by a factor of more than 105 for some neurons). (Neuron from B.B. Boycott, in Essays on the Nervous System [R. Bellairs and E.G. Gray, eds]. Oxford, UK: Clarendon Press, 1974.)
Proteins that participate Cell Growth Control 1. Growth Factors 2. Growth Factor Receptors 3. Intracellular Transducers 4. Transcription Factors 5. DNA Repair Proteins 6. Cell Cycle Control Proteins 7. Anti Death Proteins
Cell growth control through Nutritional stress These graphs show the relationship between growth rate, cell size, and cell cycle time. (A) If cell division continued at an unchanged rate when cells were starved and stopped growing, the daughter cells produced at each division would become progressively smaller. (B) Yeast cells respond to some forms of nutritional deprivation by slowing the rate of progress through the cell cycle so that the cells have more time to grow. As a result, cell size remains unchanged or is reduced slightly. (A unit of time is the cycle time observed when nutrients are in excess.)
Fresh Growth Medium Stimulates Proliferation in a confluent cell monolayer Cells in a confluent monolayer do not divide (gray). The cells resume dividing (green) when exposed directly to fresh culture medium. Apparently, in the undisturbed confluent monolayer, proliferation has halted because the medium close to the cells is depleted of mitogens, for which the cells compete
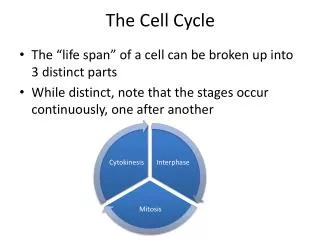
The Eukaryotic Cell Cycle In most growing cells, the four phases proceed successively, taking from 10-20 hours depending on cell type and developmental state. Interphase comprises the G1, S, and G2 phases. DNA is synthesized in S, and other cellular macromolecules are synthesized throughout interphase, so the cell roughly doubles its mass. During G2 the cell is prepared for the mitotic (M) phase, when the genetic material is evenly partitioned and the cell divides. Nondividing cells exit the normal cycle, entering the quiescent G0 state.
The Cell Cycle The cell cycle, is highly regulated in multicellular organisms: G0: Stationary or resting phase M : Mitotic phase G1: Growth phase I (cell growth and preparation for the synthetic phase) S: Synthetic phase (replication of DNA) G2: Growth phase II (Cell growth and preparation for cell division)
The fate of a single parental chromosome throughout the eukaryotic cell cycle
The control of the cell cycle The essential processes of the cell cycle such as DNA replication, mitosis, and cytokinesis are triggered by a cell-cycle control system. By analogy with a washing machine, the cell-cycle control system is shown here as a central arm the controller that rotates clockwise, triggering essential processes when it reaches specific points on the outer dial.
Checkpoints in the cell-cycle control system Information about the completion of cell-cycle events, as well as signals from the environment, can cause the control system to arrest the cycle at specific checkpoints. The most prominent checkpoints occur at locations marked with yellow boxes. 1. G1/S checkpoint 2. G2/M Checkpoint 3. G0/G1 Checkpoint
Cell Cycle Checkpoints are regulated by Cyclins and Cyclin Dependent Kinases • There are four classes of cyclins, each defined by the stage of the cell cycle at which they bind Cdks and function. Three of these classes are required in all eucaryotic cells: • G1/S-cyclins bind Cdks at the end of G1 and commit the cell to DNA replication. • S-cyclins bind Cdks during S phase and are required for the initiation of DNA replication. • M-cyclins promote the events of mitosis. • G1-cyclins, helps promote passage through Start or the restriction point in late G1.
The structural basis of Cdk activation These drawings are based on three-dimensional structures of human Cdk2, as determined by x-ray crystallography. The location of the bound ATP is indicated. The enzyme is shown in three states. (A) In the inactive state, without cyclin bound, the active site is blocked by a region of the protein called the T-loop (red). (B) The binding of cyclin causes the T-loop to move out of the active site, resulting in partial activation of the Cdk2. (C) Phosphorylation of Cdk2 (by CAK) at a threonine residue in the T-loop further activates the enzyme by changing the shape of the T-loop, improving the ability of the enzyme to bind its protein substrates.
The regulation of Cdk activity by inhibitory phosphorylation The active cyclin-Cdk complex is turned off when the kinase Wee1 phosphorylates two closely spaced sites above the active site. Removal of these phosphates by the phosphatase Cdc25 results in activation of the cyclin-Cdk complex. For simplicity, only one inhibitory phosphate is shown. The activating phosphate is added by CAK,
The inhibition of a cyclin-Cdk complex by a CK Inhibitor This drawing is based on the three-dimensional structure of the human cyclin A-Cdk2 complex bound to the CKI p27, as determined by x-ray crystallography. The p27 binds to both the cyclin and Cdk in the complex, distorting the active site of the Cdk. It also inserts into the ATP-binding site, further inhibiting the enzyme activity.
A simplified view of the core of the cell-cycle control system Cdk associates successively with different cyclins to trigger the different events of the cycle. Cdk activity is usually terminated by cyclin degradation. For simplicity, only the cyclins that act in S phase (S-cyclin) and M phase (M-cyclin) are shown, and they interact with a single Cdk; as indicated, the resulting cyclin-Cdk complexes are referred to as S-Cdk and M-Cdk, respectively.
Protein Degradation is Required for progression through Cell Cycle Passage through three critical cell-cycle transitions, G1-S phase, G2-M phase (metaphase → anaphase, and anaphase → telophase and cytokinesis), is irreversible because these transitions are triggered by the regulated degradation of proteins, an irreversible process. As a consequence, cells are forced to traverse the cell cycle in one direction only. In higher organisms, control of the cell cycle is achieved primarily by regulating the synthesis and activity of G1 Cdk complexes Extracellular growth factors, called mitogens, induce the synthesis of G1 Cdk complexes. The activity of these and other Cdk complexes is regulated by phosphorylation at specific inhibitory and activating sites in the catalytic subunit. The point in late G1 where passage through the cell cycle becomes independent of mitogens is called the restriction point
Cell Phase Specific Cyclins Activity of mammalian Cdk-cyclin complexes through the course of the cell cycle in G0 cells induced to divide by treatment with growth factors. The width of the colored bands is approximately proportional to the protein kinase activity of the indicated complexes. Cyclin D refers to all three D-type cyclins.
Regulation of mitotic cyclin B levels in M-Phase The anaphase-promoting complex (APC) is activated only when MPF activity is high. Binding of the active APC and E2 covalently linked to a ubiquitin (not shown) to the cyclin B destruction box leads to the addition of multiple ubiquitin (Ubi) molecules. As the polyubiquitinated cyclin B is degraded, MPF activity declines, triggering the onset of telophase. Following cytokinesis, synthesis of cyclin B occurs in the interphase daughter cells. APC activity remains high until late in the G1 of the next cell cycle when it is inactivated by a G1 Cdk complex. When the MPF activity rises enough, another mitoses ensues.
Growth Promoter and Growth Suppressor Genes control the Cell Cycle Growth Promoters: EGF, PDGF, Myc Growth Suppressors:Rb, P53, P16, etc.
Growth Factors Promote Cell Growth In this simplified scheme, activation of cell-surface receptors leads to the activation of PI 3-kinase, which promotes protein synthesis, at least partly through the activation of eIF4E and S6 kinase. Growth factors also inhibit protein breakdown (not shown) by poorly understood pathways
Growth Supressor Retinoblastoma (Rb) • pRB is a DNA binding protein expressed in every cell type and is an important regulator of the cell cycle. • Activated (Unphosphorylated) pRB acts as a ‘brake’ in the cell cycle at G0/G1 boundary. • During G1, Phosphorylation of pRB (by Cdks) inactivates the protein and allows cell cycle progression into S phase. G0/G1
Growth Suppressor Functions of Rb • The G0/G1 boundary is a particularly important control point because this acts as a commitment to cell division. • Cyclins (A, B, C, D) and Cyclin Dependent Kinases (Cdks) are responsible for phosphorylating and inactivating Rb. • These are in turn regulated by Cyclin dependent Kinase Inhibitors like Cip/Kip and INK4 gene products
The G0/G1 Boundry • The G0/G1 boundary is a particularly important control point because this acts as a commitment to cell division.
E2F as a growth Promoter E2F is a transcription factor that activates several replication and growth related genes. During G0/G1 transition, E2F is bound to activated or unphosphorylated RB, and as a consequence is unable to activate transcription. The RB-E2F complex can be disrupted by phosphorylation of RB or by DNA viral oncoproteins that bind RB. The release of free E2F leads to transcriptional activation of genes containing E2F binding sites, such as the adenovirus E2 gene, cmyc, c-myb, dhfr, DNA polymerase a, of the transcriptional machinery.
Mechanisms controlling S-phase initiation in animal cells G1-Cdk activity (cyclin D-Cdk4) initiates Rb phosphorylation. This inactivates Rb, freeing E2F to activate the transcription of S-phase genes, including the genes for a G1/S-cyclin (cyclin E) and S-cyclin (cyclin A). The resulting appearance of G1/S-Cdk and S-Cdk activities further enhances Rb phosphorylation, forming a positive feedback loop. E2F acts back to stimulate the transcription of its own gene, forming another positive feedback loop.
Restriction Point control Growth supressor proteins such as Rb and P16 control the cycle cycle check points. These proteins prevent excessive activation of CycD and E2F at the G1/S phase boundry.
Regulation of Rb and E2F activities in late G1 Stimulation of G0 cells with mitogens induces expression of Cdk4, Cdk6, D-type cyclins and E2F transcription factors (E2Fs), all encoded by delayed-response genes. Interaction of E2Fs with hypophosphorylated Rb protein initially inhibits E2F activity. When signaling from mitogens is sustained, the resulting Cdk4 cyclin D and Cdk6 cyclin D complexes (Cdk4/6 cyclin D) initiate the phosphorylation of Rb, converting some E2F to the active form. Active E2F then stimulates its own synthesis and the synthesis of Cdk2 and cyclin E. Cdk2 cyclin E further stimulates Rb phosphorylation releasing more E2F activity. These processes result in positive feedback loops (blue arrows) leading to a rapid rise in both E2F and Cdk2 cyclin E activity as the cell approaches the G1→S transition.
p53 • p53 has been called the ‘guardian of the genome’ as it directs cells with damaged DNA that cannot be repaired to undergo apoptosis by inducing the expression of apoptosis genes (P21, Bax, mdm-2)
Cell-Cycle Progression is Blocked by DNA Damage and p53 at the G1/S checkpoint G1/S Checkpoint
p53-induced cell-cycle arrest in response to DNA damage The normally unstable p53 protein is stabilized by damaged DNA, so its concentration increases. Acting as a transcription factor, p53 induces expression of p21CIP, a cyclin-kinase inhibitor that inhibits all Cdk1-, Cdk2-, Cdk4-, and Cdk6-cyclin complexes. Binding of p21CIP to these Cdk-cyclin complexes leads to cell cycle arrest in G1 and G2.
INK 4 P16 P14 P53 Rb Cyc D Cyc B Bax P21 E2F Bcl2 INK4 Locus (P14, P15, P16, P19)Growth Suppressors • The INK4 locus at 9p21, gives rise to multiple proteins after differential splicing. • These proteins are involved cell cycle control through Rb and P53 growth suppressors.
An overview of the cell-cycle control system The core of the cell-cycle control system consists of a series of cyclin-Cdk complexes (yellow). The activity of each complex is also influenced by various inhibitory checkpoint mechanisms, which provide information about the extracellular environment, cell damage, and incomplete cell-cycle events (top). These mechanisms are not present in all cell types; many are missing in early embryonic cell cycles, for example.