Chemiosmotic Paradigm in Microscopic Systems
E N D
Presentation Transcript
MEMBRANE POTENTIALS AND pH GRADIENTS IN MICROSCOPIC SYSTEMS:THE CHEMIOSMOTIC PARADIGMH.R. Kaback
A Little History: In 1960-61, the British scientist Peter Mitchell put forward the heretical postulate that a pH gradient (the so-called Proton-Motive Force) across the mitochondrial or bacterial membrane is the immediate driving force for oxidative phosphorylation, as well as the accumulation of metabolites against a concentration gradient (The Chemiosmotic Hypothesis). At about the same time, Robert Crane of Rutgers University suggested that sodium gradients are responsible for driving glucose accumulation across the intestinal epithelium.
In the late ‘60s, Andre Jagendorf of Cornell University carried out the first experiments that made scientists begin to take Mitchell’s hypothesis seriously. By using thylakoids (the intracellular organelles responsible for photophosphorylation in plants), Jagendorf demonstrated that sudden acidification of the external medium leads to synthesis of ATP. Over the ensuing 10 years, work in many laboratories using a variety of techniques with different experimental systems showed virtually unequivocally that Mitchell’s Chemiosmotic Hypothesis is the paradigm for bioenergetics in energy transducing membranes, and in 1977, Mitchell was awarded the Nobel Prize in Chemistry.
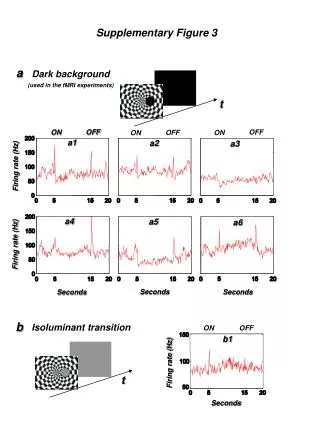
Clearly at this level, the major questions are two-fold: a. Are membrane potentials and pH gradients present in microscopic systems (too small to be impaled by microelectrodes)? b. If there are membrane potentials and pH gradients in these systems, are they of sufficient magnitude to drive the process in question?In this lecture, the focus is on right-side-out (RSO) membrane vesicles from E. coli and respiration-driven active transport in this well-defined system with the goal of providing a basic intuitive understanding of the Chemiosmotic Paradigm with respect to active transport. We will then see how the principles apply to more complex eucaryotic systems.
Although there are instances in which loops probably occur in respiratory chains, things are generally more complicated. For example, certain terminal oxidases in purified form pump protons by themselves when reconstituted into artificial membranes. There are also respiratory chains in some bacteria that pump sodium. The point is that loops in the respiratory chain provide a simple conceptual possibility for how electron transfer might be coupled to proton pumping.
From the decrease in the dialyzable [14C]acetate concentration upon addition of ASC-PMS, the quantity of acetate accumulated by the vesicles can be readily calculated per mg of vesicle protein, and knowing the intravesicular volume per mg of vesicle protein, this value can be transformed into the internal concentration of [14C]acetate (in molarity). Since the external concentration of the weak acid in the medium surrounding the vesicles is given directly from the flow dialysis profile, the concentration gradient of [14C]acetate in the presence of ASC-PMS is determined from [acetate]in/[acetate]out and converted in mV by using the constants shown in Figure 1 (2.3RT/F60 at room temperature).