Investigating Plasmid DNA Topology Changes Induced by Sequence-Specific DNA-Binding Proteins
This study explores the influence of sequence-specific DNA-binding proteins on the linking number (Lk) of plasmid DNA. We utilized two methods: ligation and Topo I, to assess plasmid behavior in the presence and absence of various proteins. Our findings demonstrated that proteins such as l.O, GalR, AraC, and LacI did not induce changes in Lk when binding sites were absent. Time-course experiments further highlighted the role of LacI in modifying topoisomerase activity. Our preliminary insights suggest a context-dependent affinity of proteins to their binding sites.


Investigating Plasmid DNA Topology Changes Induced by Sequence-Specific DNA-Binding Proteins
E N D
Presentation Transcript
SUPPLEMENTARY DATA Figure S1. The experimental strategy to determine the linking number difference of plasmids in the presence of a sequence-specific DNA-binding protein and in its absence. (A) The ligation method. (B) The Topo I (topoisomerase I) method.
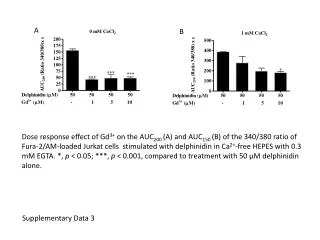
Figure S2. Protein-induced DLk or b required the presence of the DNA-binding sites in the plasmid DNA templates. Experiments to determine protein-induced DLk were performed as described under Materials and Methods. DNA topoisomers were resolved with electrophoresis in 1% agarose gel containing 0.5 mg/ml chloroquine. (A) l O protein failed to induce a DLk or b to plasmid pYZX43 that does not contain l O-binding sites (lanes 1-3). Lanes 4-6 are DNA samples from reaction mixtures using plasmid pCB37 that contains 16 l O-bindinging sites (iteron III of l DNA replication origin). In addition to 0.0625 nM of plasmid pYZX43 (lanes 1-3) or pCB37 (lanes 4-6), the reaction mixtures for the DNA samples applied to lanes 1-3 or lanes 4-6 also contains 0, 5, and 10 nM of l O protein (as a dimer), respectively. (B) GalR failed to induce a DLk or b to plasmid pYZX43 that does not contain gal operators (lanes 1-3). Lanes 4-6 are DNA samples from reaction mixture for plasmid pCB42 that contains 18 gal operators. In addition to 0.0556 nM of plasmid pYZX43 (lanes 1-3) or pCB42 (lanes 4-6), the reaction mixtures for the DNA samples applied to lanes 1-3 or lanes 4-6 also contains 0, 3, and 6 nM of GalR,
respectively. (C) AraC failed to induce a DLk or b to plasmid pYZX12 that does not contain AraC-binding sites. Lanes 4-6 are DNA samples from reaction mixture for plasmid pYZX42 that contains 14 araI sites. In addition to 0.0714 nM of plasmid pYZX12 (lanes 1-3) or pYZX42 (lanes 4-6), the reaction mixtures for the DNA samples applied to lanes 1-3 or lanes 4-6 also contains 0, 3, and 6 nM of AraC, respectively. (D) LacI failed to induce a DLk or b to plasmid pCB42 that does not contain lac operators. Lanes 4-6 are DNA samples from reaction mixture for plasmid pYZX46 that contains 19 lac O1 operators. In addition to 0.0526 nM of plasmid pCB42 (lanes 1-3) or pYZX46 (lanes 4-6), the reaction mixtures for the DNA samples applied to lanes 1-3 or lanes 4-6 also contains 0, 2, and 10 nM of LacI, respectively.
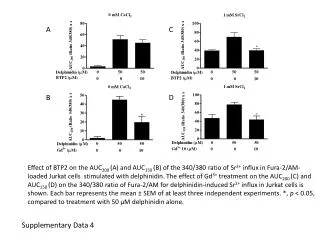
Figure S3. Time course of topoisomerase I in the presence (lanes 1 to 6) and in the absence (lanes 7 to 9) of LacI. The experiments to determine LacI-induced DLk were performed according to the Topo I method as described under Materials and Methods except for the relaxation time by topoisomerase I. Plasmid pYZX46 that contains 19 lac O1 operators was used as the DNA template. DNA topoisomers were resolved with electrophoresis in 1% agarose gel containing 0.5 mg/ml chloroquine. Lanes 1 to 6 contain DNA samples from reaction mixtures in the presence of 2 nM of LacI. Lanes 7 to 9 are DNA samples from reaction mixture in the absence of LacI.
Supplementary Discussion Comment: The method assumes that the affinity of the DNA-binding protein for each individual site is the same. Binding is often context-dependent and the authors need to justify this assumption, ideally showing some data. Our response: yes, we also agree with this general comment. Regardless, two proteins used in this paper, GalR and l O protein, bind to their recognition sites without cooperativity. For example, Brenowitz et al. demonstrated that Gal repressor binds to its DNA operators non-cooperatively under physiological conditions (Brenowitz, M., et al. (1990) Biochemistry, 29, 3374-3383). l O protein has four binding sites in the l DNA replication origin (four iterons) that have different sequences. However, O protein binds to the four iterons with the same affinity. Our preliminary results and results from Dr. Roger McMacken’s lab at Johns Hopkins University (personal communication) showed that l O protein binds to its recognition site non-cooperatively. We believe that this assumption is valid for GalR and l O protein. For other DNA-binding proteins, especially for those binding to their recognition sites cooperatively, more consideration is needed. We also ignored context-dependent differences in binding affinity.