Download

1 / 57
660 likes | 1.01k Vues
Cell biology 2014 (revised 11/2-14). Lecture 8 & 9:. The cytoskeleton Function, design and regulation. Chapter 16 965-1020 1026-1050 A lot of reading! Focus on general principles and topics highlighted in the lecture synopsis .

E N D
Cell biology 2014 (revised 11/2-14) Lecture 8 & 9: The cytoskeleton Function, design and regulation Chapter 16 965-1020 1026-1050 A lot of reading! Focus on general principles and topics highlighted in the lecture synopsis Cell Biology interactive media ”video” or ”animation”
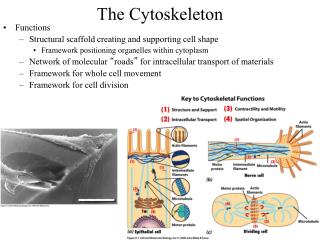
Classical cytoskeletons Microtubules Actin filaments (Microfilaments) Intermediate filaments
1. Why do we need a cytoskeleton? Cell containing cytoskeleton Cell without cytoskeleton • Establishment of cellular • shape and intracellular • organization ER Golgi 1 • Resistance against • mechanical stress 1
2. Why do we need a cytoskeleton? Cell containing cytoskeleton Cell without cytoskeleton • Cellularappendages • Cell locomotion • Genomic and • cellular division
Principle architecture of cytoskeletal filaments Actin filaments Intermediate filaments Microtubules 7 nm 10 nm 25 nm Tubulin heterodimer Actin Subunits: A family of coiled-coil proteins
Intermediate filaments – structure and function Non-polar filaments Cytosol: support of cell layers ( tensile stress ) Nucleus: supporting the nuclear envelope amphipathic a-helical monomers + Tetramer of coiled-coil dimers Cell adhesion (desmosome) animation16.4 -Intermediate _filament
Tissue specific intermediate filaments Intermediate filaments can be composed of either: Homodimers or heterodimers - Intermediate filament super-family: >60 genes in mammals Cytosolic Nuclear Protein: Cyto- keratins Vimentin, Desmin Neuro- filaments Lamins Lining of the nuclear membrane of all cells Location: Epithelia Neurons Cells in connective- and muscle tissue
P P P P P P P P P P P P P P P P P P P P P P P P Intermediate filaments in epidermis Keratin: 1 + 10 5 + 14 ECM (Basal lamina)
Actin filaments – structure and function Structure - Polar filaments composed of actin Functions - Linking the interior to the exterior ( ) - Contraction ( ) - Spreading & protrusions cell shape - Locomotion - Contractile ring during cell division ECM Video 01.1-keratocyte_dance Video 22.7 –neurite_outgrowth
Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Actin filaments are dynamic in migrating cells Rapid assembly and disassembly is central to a variety of functions, such as cell remodeling and locomotion Stimuli Time
Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Subunit interactions within actin filaments A polymer with only longitudinal subunit interactions uniform (and poor) polymer stability + + Protofilament (proto = a prefix meaning the “earliest”) A polymer with both longitudinal and lateral subunit interactions stability within the polymer but dynamic ends Internal stability Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Dynamic ends
Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Fanta Fanta Fanta Fanta Nucleation of actin filaments Spontaneous nucleation is slow because the initial interactions are unstable (low degree of cooperativity) Spontaneous nucleation Spatially regulated nucleation factors local nucleation Fanta Fanta Fanta Fanta Nucleation factor
Fanta Fanta Fanta Fanta Fanta Fanta Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Fanta Fanta Fanta Fanta Fanta Fanta Fanta Fanta Fanta Fanta Fanta Fanta Control of actin filament nucleation No availabile nucleation factor Inactive nucleation factor No (specific) nucleation No (specific) nucleation Localized activation of nucleation factors Global activation of nucleation factors Local nucleation Global nucleation
Actin nucleation factors Arp 2/3 Formin + end Arp 2/3 may also bind pre-existing filaments to create branching + end - end - end
Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Concept of the critical free concentration The monomer ( = soluble subunit) concentration ( = [Free]) at steady state is referred to as the critical concentration 0 100% Steady state Elongation Monomer concentration [Free subunits] % Subunits in filament (% Bound) Spontaneous nucleation 0 % 60 Time (minutes)
Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Dynamics at different free concentrations [Monomer] = [Free subunits] [Monomers] > critical concentration, e.g. 3 nM Net polymerization [Monomers] = critical concentration, e.g. 2 nM No net effect on polymer length [Monomers] < critical concentration, e.g. 1 nM Net depolymerization
Nucleotide turnover in cytoskeletal subunits Actin subunits bind ATP Tubulin heterodimers bind GTP The subunit changes its conformation upon nucleotide hydrolysis Subunit bound to a nucleoside triphosphate Coca Cola Nucleotide hydrolysis Subunit bound to a nucleoside diphosphate Coca Cola A nucleoside is the portion of a nucleotide that doesn't include the phosphate groups
Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola Coca Cola I. ATP fueled actin treadmilling ”Low” critical concentration (e.g. 2 nM) ”High” critical concentration (e.g. 10 nM) - end + end Treadmilling occurs when [Monomers] (i.e. [Free subunits]) is between the critical concentrations at the two ends (2 10 nM) ADP ATP During treadmilling the filament length remains constant, while subunits are added at the (+) end and dissociate from the (-) end
II. ATP fueled actin treadmilling ADP P P :interaction strength + - ADP ADP ADP ADP ATP ADP ATP P P ADP ADP ADP ATP [ADP] << [ATP] ADP ATP ATP ATP Time - + = Fluorescent actin Treadmilling actin subunits "move" towards the (-) end
Treadmilling requires actin severing [G-actin] = [Monomer] = [Free] Arp2/3 stabilizes the (-) end - + Polymerization ceases due to low [G-actin] A severing protein – ADF/Cofilin – binds to ADP-actin containing filament Polymerization at the (+) end can resume and the filament will treadmill, which will facilitate continuous growth at the (+) end ADP/ATP exchange Significance ?! slide 10 & 52
Microtubules Structure - Hollow polar tubes of tubulin protofilaments Tubulin heterodimer b-tubulin a-tubulin Protofilament Microtubule Function Exert both pushing and pulling forces Structural support and railroad tracks, which establish intracellular organization Locomotion by cellular appendages (cilia and flagella) Segregation of chromosomes during mitosis
Pushing and pulling by microtubules during mitosis Interphase (G2) Prophase Telophase/ cytokinesis Prometaphase Anaphase Metaphase Astral- video 13.2 – biosy_secret_path Video 17.7 – mitotic_spindle Overlap- Kinetochore MT
The centrosome – the site for microtubule nucleation The centrosome contains ~100 g-tubulin ring complexes, which act as nucleation sites for microtubule assembly Centriole pair g-Tubulin Ring Complex Minus-end All subunits are encoded for by the genome, but assembly requires an inherited copy as a template Plus-end
- - - - - + + + + + Different microtubules arrangements + + + - • “Most” cell types + + + + • Columnar epithelial cells • (small intestine) - + - - + + - - + + - • Neurons +
GTP GTP GTP GTP GTP GTP GTP GTP GTP GTP Pi GTP hydrolysis at the E-site of the tubulin heterodimer E-site E-site E-site = Exchangeable site b a GTP GDP b a Catalytic loop Catalytic loop
Proteins that control microtubule dynamics Stabilization by Microtubule Associated Proteins (MAPs) Multivalent binding along the polymer Destabilization by catastrophe promoters Peeling of proto-filaments at the end
GTP-tubulin GDP-tubulin MT dynamics – catastrophe GTP cap (E-site exposed) Catastrophe promoting protein + end (delay in GTP hydrolysis) • end • (nucleated at • the centrosome) The + end is “capped” by GTP-tubulin Catastrophe, followed by depolymerization Peeling of proto-filament
GTP-tubulin GDP-tubulin MT dynamics – rescue Paus Depolymerization Regain of GTP cap through re-initiated polymerization : Rescue promoting protein video 16.1- MT_instability
Dynamic instability – stochastic switches [GDP] << [GTP] GTP GDP + end Polymerization Depolymerization Catastrophe Rescue • end • (nucleated at • the centrosom) : Rescue promoting protein : Catastrophe promoting protein Dynamic instability ”search and capture” of a variety of structures
P Cdk Cdk M M P P Cell cycle regulation of microtubule dynamics Interphase Mitosis (active Cdk/M) P P Few and long microtubules: - Few nucleation events - Slow dynamics Many and short microtubules: - Many nucleation events (5x) - Rapid dynamics (10x) video 16.5- microtubule_dynamics Note- visualization by fusion to a fluorescent protein (EB1-GFP& aTub-GFP)
Capture of kinetochores by microtubules MTs continuously searches the cellular space... 1 ...and are stabilized at the kinetochores of chromosomes 2 * * * * * * * * * * Finally, both kinetochores are captured by MTs from opposite centrosomes. *The sister chromatid pair is positioned at the cellular equator by the polar ejection force generated by MTs (* ) 3 Generation of pulling force
Unidirectional transport on polar polymers Candy Check-out Polarity in a queue at the supermarket Motor proteins (unidirectional movement) + end - end
A B A A B A B B A B Movement of MT dependent motor proteins Dynein Kinesin - end + end Head-over-head walking (an ATP dependent process) 1. 2. 3. 4. 5. animation 16.7- kinesin
+ + MT dependentpushingforcesduringmitosis Kinesin dimer Kinesin dependent pushing forces via anti-parallel MTs are required for: Prophase Anaphase (B)
Control of division plane in epithelia Incorrect Correct
Control of division plane by astral microtubules Dynamic (astral) microtubules are stabilized by tip binding proteins ( ) at specific sites at the cell cortex 1. Membrane anchored dynein ( ) pulls at astral microtubules - 2. 1. + 2. Basal lamina(ECM) Pulling forces specify the correct division plane 3. 3.
Cell polarization by localized MT stabilization A non-polarized cell in which MTs search the intracellular space Stabilization of MTs that encounters localized tip-binding proteins ( ) Reorientation of the MT- system by membrane anchored dyneins ( ) A polarized cell: stabilized MTs serve as rail tracks that transport membrane vesicles and actin regulatory proteins to MT (+) ends
MT-dependent trafficking in the cell + Kinesin Dynein Virus ER + - - Vesicle Axon Golgi Lysosome Synapse Endocytosis Mitochondrion Exocytosis video 16.6- organelle_movement
Cellular appendages built of microtubules Cilia Flagella • - 5 -10 mm appendages projecting • from cell surfaces • - Capable of movement • - Moves fluids over • the cell surface - In essence a cilia, but longer (100-200 mm) - Only one per cell - Move the cell in a wavelike fashion
Arrangement of microtubules in cilia and flagella Flagella Axoneme Cilia Basal body Axoneme: the part of a cilia or flagella that bends back and fourth
The beating of a cilia The beating of cilia is dependent on MT bending forces Power Stroke (energi input) Axoneme Basal body Recovery Stroke (back to default)
1. 2. 3. Dynein dependent MT bending in cilia and flagella Nexin, holds the MTs together Anchorage to dynein tail Bending of MTs upon dynein movement
+ Head ADP +Pi - Myosin: a family of (+) end-directed actin motors Myosin bound to actin filament ATP binding dissociates myosin ATP ATP is rapidly hydrolyzed, which cause a simultaneous conformational change ADP +Pi Following ATP hydrolysis, myosin binds an actin subunit Binding to actin causes the release of ADP + Pi. This results in the conformational change termed the “power stroke” ADP +Pi Video 16.9 –crawling_actin
+ - - - - + “Non-muscular” myosin family members A large family of related (+) end directed motors. Example of functions: + Monomeric myosin Transport(short range) Non-muscular myosin II Contraction (movement towards the +ends of two anti-parallel actin filaments) + Cargo
Lumen Lumen Muscles – a brief overview • Skeletal muscle, fused myoblast that forms a multinucleated cell • (fast but non-persistent) • Cardiac muscle cells • (persistent) • Smooth muscle cells • i) surrounds hollow organs – intestines and blood vessels • ii) Arrectorpili muscles attached to hair follicles • (slow and very persistent) video 16.11- beating_heart
Principle of skeletal muscle contraction When stimulated to contract, the heads of the bipolar myosin filament walk along actin in repeated cycles of attachment and detachment contraction of the sarcomere unit Actin Myosin Actin Myosin Contraction + + + + Sarcomere Sarcomere • - The actin and myosin filaments remain the same length • - The sarcomere length shortens because the actin and myosin filaments slide relative each other animation 16.8- myosin (compare with picture 43)
Tropomyosin Tropomyosin Tropomyosin Tropomyosin Tropomyosin Tropomyosin Tropomyosin Tropomyosin Regulation of skeletal muscle contraction • Tropomyosin binds along the actin filament: • No contact between actin and myosin filaments Contraction is initiated by an increase of cytosolic Ca2+: Troponin mediated translocation of tropomyosin Ca2+ animation 16.10- muscle_contraction
- - - - Higher-order architecture of actin filaments Actin filaments (in non-muscle cells) may associate into bundles or networks via different cross-linking proteins Anti-parallel bundles allowing access to myosin II Sparse 3D network Tight parallel bundles + + a-actinin a-actinin Fimbrin Fimbrin + + Filamin short and thin fibers Long and thick fibers
Cell migration requires locally acting GTP switches Rho Rac Cdc42 GTP GTP GTP - - + - - + Stress fibers (contraction) Actin web (tread milling) Actin bundles (protrusions) + Rho/Rac/Cdc42 are GTP switches (similar to Ras) +
An integrated view of actin dependent migration Actin structure: Stress fibers (contractile) Lamellipodia (pseudopodia) Filopodia (microspikes) + - - - + Chemoattractant (e.g. PDGF) + + - Rho family member: Rho Rac Cdc42 video 23.9- wound_healing