Download

1 / 100
1.05k likes | 1.55k Vues
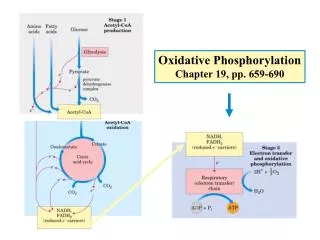
Chapter 19 Oxidative phosphorylation and photophosphorylation. Electron transferring via a chain of membrane bound carriers, across membrane proton gradient, ATP synthesis (with O 2 consumed or produced). Oxidative phosphorylation occurs on the convoluted inner membrane of mitochondria

E N D
Chapter 19 Oxidative phosphorylation and photophosphorylation Electron transferring via a chain of membrane bound carriers, across membrane proton gradient, ATP synthesis (with O2 consumed or produced)
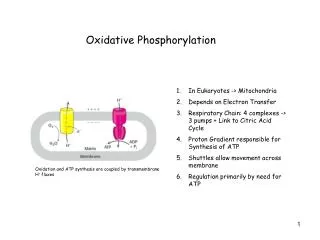
Oxidative phosphorylation occurs on the convolutedinner membrane of mitochondria (cristae) (or plasma membrane In bacteria)
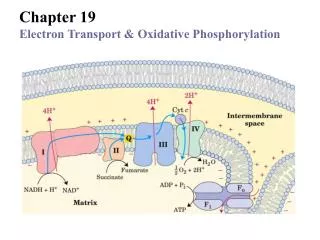
E`0 = -0.320 V A 1.14-volt potential difference (E`0) between NADH and O2 drives electron flow through the respiratory chain I II III The respiratory chain consists of 4 multi- subunit complexes and two mobile e- carriers. IV E`0 = 0.816 V
Various prosthetic groups act as intermediate electron carriers in the respiratory chain
Q is a mobile one/two e-/H+ carrier The only e- carrier not bound to a protein Q/QH2 diffuses freely in the lipid bilayer
Also found in Hemoglobin & myoglobin Bound to cytochromes tightly and covalently. Heme groups of cytochrome proteins Bound to cytochromes tightly but noncovalently Three types of heme groups are found in cytochromes.
Reduced cytochromes has three absorption bands in the visible wavelengths c b a The reduced (Fe2+) state of cytochromes a, b, and c has a longest wavelength band near 600, 560, and 550 nm respectively.
2Fe-2S 4Fe-4S At least eight different types of iron-sulfur centers act in the respiratory chain: iron atoms cycle between Fe2+ (reduced) and Fe3+ (oxidized).
Electrons of NADH and FADH2 are transferred to O2 via many intermediate electron carriers on the way.
NADH enters at NADH dehydrogenase (also NADH:ubiquinone oxidoreductase or NADH-Q oxidoreductase) Consisting at least 34 polypeptide chains “L” shaped, ~880 KDa (ribosome: ~2500 KDa) exergonic endergonic The coupling mechanism between e- transferring and H+ pumping is still unknown
succinate Dehydrogenase (complex II) FADH2 of flavoproteins also transfer their electrons to ubiquinone (Q), with no H+ pumped.
The three core subunits Cytochrome bc1 complex (Complex III) passes 2e- from 1 QH2 to 2 cytochrome c Consists of two 11-subunit monomer
e- transferring & H+ pumping in Complex III via the Q cycle (for each 2 e- transferred, 4H+ are released at the P side and 2H+ taken from the N side) The Q cycle The Q cycle Qp Qp QN QN 1st QH2 2nd QH2 A switch between 2e- carrier (QH2) and 1e- carrier (Cyt c)
3 subunits (out of the 13) seem to be critical CuA-CuA CuB Copper center A Heme a CuA Heme a3 CuA Cytochrome c oxidase (Complex IV ), the terminal enzyme of the respiratory chain, contains 3 Cu and 2 heme A groups as electron carriers
4e- are transferred from 4 Cyt c to 1 O2 to make 2H2O in Complex IV, with 4H+ taken from the matrix to make 2H2O& 4H+ pumped out. ?
A proposed reaction cycle for the four-electron reduction of O2 by cytochrome c oxidase (at the Heme a3-CuB center) Oxidized Fe-Cu center Reduced Fe-Cu center 2e- are transferred to the bound O2 Reduced a3 binds O2
~ 10 protons are pumped per NADH, and ~6 per FADH2 oxidized via the respiratory chain. A transmembrane H+ gradient is generated using energy released from the electron transferring: the electron motive force is converted to an proton motive force.
The order of the many electron carriers on the respiratory chain have been elucidated via various studies
Electron carriers may have an order of increasing E`0 (in complex I) (in complex III) (in complex III) (in complex IV) (in complex IV)
Oxidation kinetics studies: Full reduction followed by sudden O2 introduction; earlier oxidation, closer to the end of the chain. Oxidized Reduced Oxidation status of the cytochromes can be followed using rapid and sensitive spectrophotometric techniques. Reduced Oxidized Reduced Studies with specific inhibitors help to reveal the orders of the e- carriers
The functional complexes have been separated and characterized
Electron transfer to O2 was found to be obligatorily coupled to ATP synthesis from ADP + Pi in isolated mitochondria
ATP is not synthesized when only ADP + Pi are added Both e- flow & ATP synthesis were suppressed When an inhibitor of respiratory chain was added O2 consumption was also not observed in the presence of inhibitors of ATP synthase e- flow and ATP synthesis were both detected when a reductant is added, e- flow was neither observed if ADP was not added to the suspension, although a reductant is provided! Electron transfer and ATP synthesis in isolated mitochondria suspensions: neither occurs without the other. O2 consumption is an indication of electron flow
How is oxidation of NADH/FADH2 coupled to the phosphorylation of ADP?
It was widely believed that ATP synthesis occurs by chemical coupling • High energy intermediates similar to 1,3-biphosphoglycerate (which is used for substrate-level phosphorylation) was once proposed to be produced first from the electron flows on both the mitochondrial and chloroplast membranes. • Phophorylated protein intermediates (as formed in the action of phosphoglycerate mutase and phosphoglucomutase) were also hypothesized. • But neither were ever revealed despite intense efforts by many investigators over several decades.
The chemiosmotic model was proposed by Peter Mitchell in 1961 to explain the coupling of electron flow and ATP synthesis
The chemiosmotic model of Mitchell: e- flow and ATP synthesis are separate events, coupled via a transmembrane H+ gradient!
Supporting evidences for the chemiosmotic coupling were collected • A closed membrane system is essential for ATP synthesis but not for the electron flow (tested with detergent or physical shearing).
Hydrophobic weak acids (e.g., DNP), uncouples ATP synthesis from electron flow. electron flow continues ATP not synthesized
proton gradient is dissipated
Dectection of transmembrane proton pumping using a pH meter
Artificially imposed proton gradient alone (in the absence of an oxidizable substrate) was found to be able to drive ATP synthesis!
The chemiosmotic theory unified the apparently disparate energy transduction processes as oxidative phosphorylation, photophosphorylation, active transport across membrane and the motion of bacterial flagella.
ATP synthase was first identified by dissociation and reconstitution studies
Abundant knob-like protruding structures were observed on the matrix side of the inner mitochondrial membrane by EM (Racker in 1960).
Reconstitution studies indicate that the “knobs” are essential for ATP synthesis the protruding F1 part catalyzes ATP synthesis/hydrolysis
The 18O exchange experiment: the G`0 for ATP synthesis on purified F1 is close to zero! (Paul Boyer, ) ATP formation/hydrolysis are readily reversible in the active site of purified F1
Measurement of Kd values: ATP has a much higher affinity than ADP to the enzyme (10 -12 M vs 10–5 M). The proton-motive force is proposed to release ATP from ATP synthase
ATP synthase, “a splendid molecular machine”, comprises a proton channel (Fo) and an ATPase (F1) F1: a3b3gde. Fo: ab2c10-12
The binding-change model was proposed to explain the action mechanism of ATP synthase • The model was proposed based on kinetic and binding studies (before 3-D structure determined). • Downhill proton movement through Fo is proposed to drive the rotation of the c-subunit ring and the asymmetrical g subunits, which will cause each of the three b subunits to interconvert between the three conformations, as a result, each of them take turns to take up ADP + Pi, synthesize ATP, and release ATP.
The estimation of H+ consumption for each ATP formed is 4 • The chemiosmotic coupling allows nonintegral stoichiometries of O2 consumption (or NADH and FADH2 oxidation) and ATP synthesis.
Binding-change mechanism of Paul Boyer (1977) Rotation of g changes lead to sequential binding changes for each b subunit. g g g
The binding-change model was elegantly supported by two other experimental observations • X-ray crystallography revealed that the three b subunits of F1 assumes three different conformations, with bound ADP, ATP analog, or empty respectively. • Rotations of the g subunit and the c subunits of the F1 unit in three discrete steps of 120o (powered by ATP hydrolysis catalyzed by the b subunits) have been directly observed using fluorescence microscopy by Dr. Kazuhiko Kinosita in 1997.
App(NH)p ADP Empty Rod-shaped g subunit. Structure determination indicates that indeed each b subunit of an ATP synthase assumes a different conformation! (John Walker, 1994)
The world’s smallest molecular motor: rotational catalysis. Fluorescencetly labeled Rotation of the g subunit and the ring of c subunits in the FoF1 complex was observed by in vitro studies using fluorescence Microscopy (K. Kinosita, 1997)
Rotation of the g subunit and the ring of c subunits in in the FoF1 complex as directly observed by in vitro studies using fluorescence microscopy: they rotate in 120-degree increments,with each step consume one ATP; operation efficiency near 100%.
Model of the E. coli ATP synthase: the proton gradient drives the rotation of the c ring using two half-channels on the a subunit. (3ATP made per 10-14 protons transported: ~3H+/ATP.)