Download

1 / 36
360 likes | 592 Vues
ATP as a Signaling Molecule. CONTENTS. What is ATP..? ATP as a Signaling Molecule ATP Storage and Release Purinergic Receptors ATP in Extracellular Signaling ATP in Intracellular Signaling Ectonucleotidases.

E N D
CONTENTS • What is ATP..? • ATP as a Signaling Molecule • ATP Storage and Release • Purinergic Receptors • ATP in Extracellular Signaling • ATP in Intracellular Signaling • Ectonucleotidases
In times of plenty, need, or stress, cells can release ATP and other nucleotides into their surroundings. • This ATP can potentially, via specific purinergic receptors, influence the ATP-releasing cells or neighboring cells by the processes of • neural transmission, • paracrine signaling, and • autocrine signaling
Physiological and Pathophysiological processes associated with purinergic receptors • synaptic transmission, • pain and touch perception, • vasomotor responses, • platelet aggregation, • endothelial release of vasorelaxants, • immune defense, • cell volume regulation, • cell proliferation and mitogenesis, • apoptosis, and epithelial ion and water transport
Action of ATP on cells depends on • storage and release of ATP, • stimulus for the release, • population of the purinergic receptors on cells, • breakdown of ATP by ectonucleotidases, • cellular signaling pathways, and • effector processes within the stimulated cells
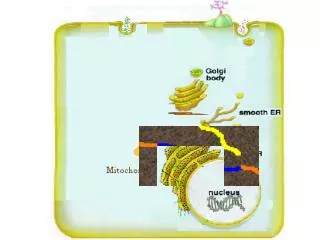
ATP storage and release • Newly synthesized ATP is first made and transported out of the mitochondrion via oxidative phosphorylation. • Steady-state cytosolic ATP conc. ~3–10 mM • Higher concentrations of ATP (up to 100s of mmol/l) are stored in secretory vesicles of neurons and released with acetylcholine, noradrenaline, and other transmitters, including VIP and neuropeptide Y.
In addition • chromaffin granules of the adrenal medulla, • serotonergic granules of platelets, and • insulin-containing granules of pancreatic-cells also store significant amounts of ATP. • ATP release via exocytosis or • Shear stress, stretch, hypoxia, inflammation, osmotic swelling, cell death, cholinergic stimulation.
Possible ATP release mechanisms in epithelia ATP-permeable release channels • Anion channels like PM Voltage Gated Anion Channels (VDAC) • Resides in apical & basolateral membranes • It was proposed that the cystic fibrosis (CF) transmembrane regulator (CFTR) or other members of the ATP-binding cassette protein family could transport ATP directly.
The existence of ion channels, that are permeated by ATP anions, was underscored by many laboratories investigating whether CFTR conducted ATP itself in addition to Clˉ or regulated a separate ion channel that was permeable to ATPˉ. • One group suggested that CFTR and the multidrug resistance transporter (mdr or P-glycoprotein) conducted ATP • Another study proposed CFTR could control maxi anion channels similar to mitochondrial porins and/or the outward rectifier Clˉ channel, either of which could transport ATP.
Connexin hemichannel • One attractive proposal is the connexin hemichannel, which, apart from being a building block for the gap junctions, is implicated in ATP release. • The hemichannels could themselves transport ATP or be associated with the cytoskeletal organization and movement of intracellular vesicles. Adenine nucleotide transporters • Adenine nucleotide transporters may also exist that may be carriers or permeases
Nucleotide/Nucleoside exchangers • Exchangers may exist for ATP in the plasma membrane, as they do in mitochondrial inner membrane, where ADP may be the exchanged substrate. Other substrates such as Na+ or Clˉ, which have favorable entry gradients, may be exchanged for ATP. ATP-filled vesicle • ATP-filled vesicles, which may also contain additional agonists or co-agonists, may fuse with the plasma membrane releasing ATP.
A high content of ATP is found in synaptic vesicles with other neurotransmitters, mast cell granules with histamine, and chromaffin granules with epinephrine. • ATP and its metabolites are known co-transmitters that modulate the effect of excitatory or inhibitory neurotransmitters. • ADP and ATP are also released by platelets on their own via exocytosis for a ‘‘self-aggregation’’ signal at the clotting zone.
Purinergic receptors • Purinoceptors, are a family of newly characterized plasma membrane molecules involved in several cellular functions such as vascular reactivity, apoptosis and cytokine secretion. • Purinoceptors are divided into two classes P1 or adenosine receptors P2, which recognize primarily extracellular ATP, ADP, UTP and UDP.
Two subclasses of P2 receptors- P2X and P2Y P2X receptors • P2X receptors are extracellular ATP-gated calcium-permeable non-selective cation channels that are modulated by extracellular Ca2+, Na+, Mg2+, H+ and metal ions, such Zn2+ and/or Cu2+. • To date, seven separate genes coding for P2X subunits have been identified, and named to as P2X1 through P2X7 • Two transmembrane domains connected by a large extracellular loop and a COOH terminal of variable length
P2Y receptors • P2Y receptors (with exception of P2Y12 and P2Y13) couple to heterotrimeric G proteins and phospholipases (primarily phospholipase Cβ) to raise intracellular free calcium concentration. • To date, 12 P2Y receptors have been cloned in humans: P2Y1, P2Y2, P2Y4, P2Y5, P2Y6, P2Y8, P2Y9 (present in NCBI as GPR23), P2Y10, P2Y11, P2Y12, P2Y13 and P2Y14. • P2Y receptors are present in almost all human tissues where they exert various biological functions based on their G-protein coupling.
All genuine P2Y receptors (except P2Y12) activate phospholipase C (PLC), phosphoinositide hydrolysis, mobilization of intracellular Ca2+, and activation of PKC • Regarding their selectivity, P2Y1, P2Y11, P2Y12, and P2Y13 are purinoceptors, P2Y6 is a pyrimidinoceptor, and P2Y2 and P2Y4 receptors have mixed selectivity. • P2Y11 stimulates PLC and adenylate cyclase, whereas P2Y12 and P2Y13 inhibit adenylate cyclase.
Other P2Y receptors can also couple to several distinct G proteins, e.g., P2Y1 and P2Y2 via Gs stimulate PLC, but via Gi they inhibit adenylate cyclase. • A multitude of receptors and signaling pathways offers a rich possibility for a cross-talk mechanism, including various isoforms of adenylyl cyclases (activated by βγ–subunits released from G proteins), Ca2+-calmodulin and/or PKC, and adenosine receptors after ATP is split into adenosine by ectonucleotidases and 5'-nucleotidase.
New pathways that may be involved in long-term regulation for non-epithelial cells metabolism cell adhesion gene expression and growth • For example, In astrocytes UTP/ATP can stimulate a mitogen-activated protein kinase (MAPK) cascade, such as extracelluar signal-regulated kinase (ERK), which involves Ca2+-independent PKC.
Topology of P2X and P2Y receptors Mammalian P2Y receptors are 328 to 370 amino acids long, transverse the membrane 7 times, and activate 1 or more G proteins. Mammalian P2X receptor monomers are 339 to 595 amino acids long and contain a long COOH terminal and a large extracellular loop, with disulfide bonds and several N-linked glycosylation sites.
Adenosine receptors • All adenosine receptors were shown to activate at least one subfamily of mitogen-activated protein kinases. • The actions of adenosine are often antagonistic or synergistic to the actions of ATP. • In the CNS, adenosine has multiple functions, such as modulation of neural development, neuron and glial signalling and the control of innate and adaptive immune systems
ATP in extracellular signaling • Once released, ATP is free to bind to P2X and P2Y receptors, on the same cell or neighboring cells. • Alternatively, ADP, a metabolite, may also bind with high affinity to a subset of P2 receptors. • UTP and UDP follow similar chemistry and have their specific P2Y receptor subtypes. • The final metabolite of ATP, adenosine, is also biologically active and binds to P1 receptors. • Once these receptors are activated, signal transduction begins and affects cell or tissue function.
ECTONUCLEOTIDASES • Once ATP leaves the cell, it is also degraded rapidly; thus, it is thought of as a local mediator that acts in an autocrine or paracrine manner within tissues and tissue microenvironments to stimulate its receptors before it is chemically modified. • Ecto-enzymes secreted into the extracellular milieu as a secreted protein or membrane-bound as an ecto-enzyme target ATP.
Two families of ectonucleotidases • E-NTPDasesare ecto-nucleoside triphosphate diphosphohydrolases that hydrolyze nucleoside 5'-tri- and diphosphates. • E-NPP, are ectonucleotide pyrophosphatase/ phosphodiesterases with a broad substrate specificity. • These can hydrolyze phosphodiester bonds of nucleotides and nucleic acids and pyrophosphatase bonds of nucleotides and nucleotide sugars. • e.g., cleavage of ATP to AMP and PPi and conversion of cAMP to AMP
Resynthesis of ATP • Secreted or membrane-bound ecto-kinases such as adenylate kinase or nucleoside monophospho- and diphosphokinases can phosphorylate nucleosides to remake 5'-AMP, ADP and, ultimately, the triphosphate nucleotide ATP.
Intracellular ATP signaling • ATP is critical in signal transduction processes. • It is used by kinases as the source of phosphate groups in their phosphate transfer reactions. • Kinase activity on substrates such as proteins or membrane lipids are a common form of signal transduction. Phosphorylation of a protein by kinase Mitogen-activated protein kinase cascade
ATP is also used by adenylate cyclase and is transformed to the second messenger molecule cyclic AMP, which is involved in triggering calcium signals by the release of calcium from intracellular stores. • This form of signal transduction is particularly important in brain function, although it is involved in the regulation of a multitude of other cellular processes.
A signal-transduction pathway for ATP release from erythrocytes • ATP is released from erythrocytes in response to a clearly defined physiological stimulus i.e reduced hemoglobin oxygen content • Suggests that signaling mechanism that relates this stimulus to ATP release must be present within that cell. • This pathway includes the heterotrimeric G protein Gi , adenylyl cyclase (AC) , protein kinase A and the cystic fibrosis transmembrane regulator (CFTR)
Proposed pathway for regulated ATP release from erythrocytes in response to passage of these cells through areas of increased oxygen demand in skeletal muscle
The increase in oxygen demand Oxygen release from hemoglobin within the erythrocyte. Decrease in Hemoglobin oxygen content Activation of the heterotrimeric G protein, Gi and release of ATP ATP released from the erythrocyte can bind to purinergic receptors (P2y) on the vascular endothelium Release of vasodilators and, an increase in blood flow (oxygen delivery)