Structure Determination
Structure Determination. X-ray and NMR. Topic 7. Chapter 4 & 5, Du and Bourne “Structural Bioinformatics”. Experimental methods. X-Ray and NMR, some history.


Structure Determination
E N D
Presentation Transcript
Structure Determination X-ray and NMR Topic 7 Chapter 4 & 5, Du and Bourne “Structural Bioinformatics”
X-Ray and NMR, some history Kendrew, J. C.; G. Bodo, H. M. Dintzis, R. G. Parrish, H. Wyckoff, D. C. Phillips (1958). "A Three-Dimensional Model of the Myoglobin Molecule Obtained by X-Ray Analysis". Nature 181 (4610): 662–666 Max Perutz and Sir John Kendrew won the Nobel Prize in Chemistry in 1962. The first de novo atomic resolution NMR structure of a globular protein, the bull seminal protease inhibitor (BUSI) was determined in 1984 by Kurt Wüthrich’s group. “Williamson, M.P., Havel, T.F. & Wüthrich, K. J. Mol. Biol. 182, 295–315 (1985).” Kurt Wüthrich was award the Nobel Prize in Chemistry in 2002 “for his development of nuclear magnetic resonance spectroscopy for determining the three-dimensional structure of biological macromolecules in solution”. Kurt Wüthrich, “The way to NMR structures of proteins”. Nature Structural Biology, 8(11):923-925, 2001
Some History about NMR by Kurt Wüthrich “The completion of the first protein NMR structure brought new, unexpected challenges. When I presented the structure of BUSI (Fig. a) in some lectures in the spring of 1984, the reaction was one of disbelief, and because of the close coincidence (Fig. b) with results from an independent crystallographic study of the homologous protein PSTI (porcine pancreatic secretorytrypsin inhibitor) it was suggested that our structure must have been modeled after this crystal structure. In a discussion following a seminar in Munich in1984, Robert Huber (Nobel Prize in Chemistry, 1988) proposed that we settle the matter by independently solving a new protein structure by X-ray crystallography and by NMR (α-amylase inhibitor tendamistat). Virtually identical three-dimensional structures of tendamistat were obtained in our laboratory by NMR in solution and in Robert Huber’s laboratory by X-ray diffraction in single crystals.” 65 amino acids Kurt Wüthrich, “The way to NMR structures of proteins”. Nature Structural Biology, 8(11):923-925, 2001
Steps in Structure Determination using X-ray Crystallography Protein purification • Grow crystals of the protein • --can take weeks to years • --not every protein can be crystallized! • Obtain the X-ray diffraction data. • Compute electron density maps. • Atomic model calculation and refinement.
Protein Crystals and Crystallization Process • Look like normal crystals, but are actually more like gels (20 to 80% solvent).
Protein Crystals and Crystallization Process Unit cell crystal
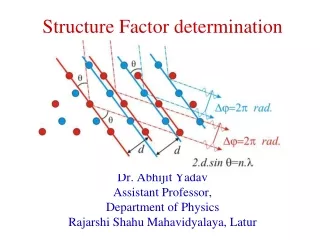
X-ray Diffraction Bragg’s law explains why cleavage faces of crystals reflect x-ray beans at certain angles of incidence (diffraction) • d = spacing between molecules in the lattice • = angle of observed diffraction • = wavelength of x-rays
Resolution is directly proportional to q In x-ray crystallography, the phrase “2.0 Å model” means that the analysis included reflections out to a distance of 1/(2.0 Å) from the center.
Resolution of X-ray Structures 3.0 Å 2.0 Å 1.0 Å Resolution is a measure of the level of detail present in the diffraction pattern and the level of detail that will be seen when the electron density map is calculated.
Phase Problem in X-ray Diffraction • Light detectors, such as photographic plates or CCDs, measure only the intensity of the light that hits them. This measurement is incomplete because a light wave has not only an amplitude (related to intensity), but also a phase, which is systematically lost in a measurement. • The phase part of the wave often contains valuable information on the sample. In x-ray crystallography, the diffraction data when properly assembled gives the amplitude of the 3D Fourier transform of the molecule's electron density in the unit cell. If the phases are known, the electron density can be simply obtained by Fourier synthesis. • There are several different methods for determining the phases: • -- Direct methods (typically not applicable to proteins) • -- Molecular replacement (based on comparisons to homologous protein w/ solved structure) • -- Isomorphous Replacement (based on comparison of two crystals, normal and one co-crystalized with heavy atom, which is not expected to affect the unit cell, thus ‘isomorphic’) • -- Anomalous Dispersion (based on single crystal, selenomethionine)
Why Solving the Phase Problem is Important? Fourier transform Fourier transform A B Mix of the magnitudes of B and the phase of A Mix of the magnitudes of A and the phase of B
Analysis of Diffraction Pattern • The diffraction pattern is analyzed by mathematical and computation methods (Fourier transform analysis) to produce an electron density map. • Note the objective result of a crystallographic experiment is not really a picture of the atoms, but a map of the distribution of electrons in the molecule, i.e. an electron density map—the x-rays are scattered from the electron cloud of the atoms. • Since the electrons are mostly tightly localized around the nuclei, the electron density map gives us a pretty good picture of the molecule.
From electron density to model Refinement Note: While some manual fitting still occurs, this process is largely done automatically by a few different computer programs.
R-factor of X-ray Structures An initial atomic model can be built by fitting the atoms to the density map. The overall model can be improved a great deal by an iterative process called refinement, in which the atomic model is adjusted to improve the agreement with the observed diffraction data. The agreement between the diffraction data and the model is measured by R-factor: F: structural factor • The R-factor is usually reported in %, i.e. an R-factor of 0.20 is reported as 20%. • R-values range from 0 (perfect fit) to 0.63 (random). • As a rule of thumb, structures with a resolution at or better than 2.5 A with R-factors below 25% tend to be largely correct. • R-free: about 10% of the observations are removed from the data set before refinement. Then, refinement is performed using the remaining 90%. The R-free value is calculated to see how well the model predicts the 10% that were not used in refinement. • ******the R-free value is a less biased.
B-factor of X-ray Structures The B-factor (or temperature factor) is an indicator of thermal motion about an atom. Note that the B-factor is a mix of real thermal displacement, static disorder (multiple but defined conformations or crystal heterogeneity) and dynamic disorder (no defined conformation), and all the overlap between these definitions. Cytochrome b5 (PDB 3B5C) highest B-factors: red lowest B-factors: blue.
X-ray crystallography and membrane proteins Intrinsic membrane proteins remain challenging to crystallize because they require detergents or other means to solubilize them in isolation, and such detergents will frequently interfere with crystallization. However, such membrane proteins are a large component of the genome and include many proteins of great physiological importance, so solving their structure remains an important open problem. Amphipathic detergent molecule Micelle From membrane integral to protein/detergent (and later crystal lattice)
Potassium channel K+ Na+ vs. Q: How can the channel allow K+ flux, but prevent Na+?
Structure Determination by NMR • Based on magnetic moments of atomic nuclei. • NMR measures the interactions of atomic nuclei. • No crystals are needed for NMR—solution-based. • Size limitations: only works well for smaller proteins (<50kD). • A typical NMR structure includes an ensemble of protein structures, all of which are consistent with the observed list of experimental restraints.
The Basics of NMR When molecules are placed in a strong magnetic field, the magnetic moments of the nuclei align with the field. Therein, each nucleus behaves like a bar magnet.
The Basics of NMR • The equilibrium alignment can be changed to an excited state by applying radio frequency (RF) pulses. • When the nuclei revert to the equilibrium they emit RF radiation that can be detected.
Chemical Shifts and J-Couplings • The chemical is the most basic of measurements in NMR. • -- The frequency of a signal is known as its chemical shift. • -- The extent of the chemical shift is related to local environment. • -- The frequency is determined relative to a reference signal. • J-coupling is the coupling between two nuclear spins due to the influence of bonding electrons on the magnetic field running between the two nuclei.
Output of a typical NMR Experiment • Nuclear Overhauser effect (NOE): inter-nuclear distances • J-Coupling: torsion angles • Chemical shift: local nuclear environment A series of protein structure models that satisfy as many of the restraints as possible are built. In the PDB, there are typically two types of coordinate entries for NMR structures. --The first includes the full ensemble from the structural determination, with each structure designated as a separate model. --The second type of entry is a minimized average structure.
NMR and X-Ray Structure Comparison “Comparison of X-ray and NMR structures: Is there a systematic difference in residue contacts between X-ray- and NMR-resolved protein structures?” Garbuzynskiy et al. Proteins, 60(1):139-147, 2005 --X-ray and NMR structures of the same protein have more differences than various X-ray structures, and even more than various NMR structures of the protein. --X-ray and NMR structures of 18 of these 78 proteins have obvious large-scale structural differences that seem to reflect a difference of crystal and solution structures. --60 pairs of structures have only small-scale differences comparable with differences between various X-ray or various NMR structures of a protein
NMR vs. X-Ray • X-ray crystallography • --need crystals • --in principal no size limit • --based on the scattering of the electron cloud of the atoms • --Quality: resolution, R-factor • Nuclear Magnetic Resonance spectroscopy (NMR) • --solution-based • --size limitation (< 50K) • --one or more models • --good for studying protein dynamics • --“resolution”: root-mean-square-deviation (RMSD) For both X-ray and NMR structures, what sets the resolution limit is the intrinsic order (or disorder) of the protein and the amount of data. The greater the amount of data, the higher the resolution
Cryo-electron microscopy (cryo-EM) ribosome rotavirus Ab fibrils