Cytoskeleton and Cell Movement
1.21k likes | 2.96k Vues
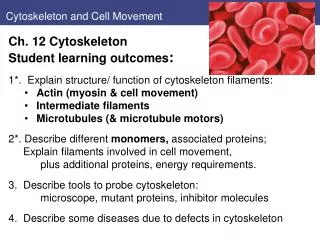
Cytoskeleton and Cell Movement. Ch. 12 Cytoskeleton Student learning outcomes : 1*. Explain structure/ function of cytoskeleton filaments: Actin (myosin & cell movement) I ntermediate filaments Microtubules (& microtubule motors)

Cytoskeleton and Cell Movement
E N D
Presentation Transcript
Cytoskeleton and Cell Movement • Ch. 12 Cytoskeleton • Student learning outcomes: • 1*. Explain structure/ function of cytoskeleton filaments: • Actin (myosin & cell movement) • Intermediate filaments • Microtubules (& microtubule motors) 2*. Describe different monomers, associated proteins; Explain filaments involved in cell movement, • plus additional proteins, energy requirements. • 3. Describe tools to probe cytoskeleton: • microscope, mutant proteins, inhibitor molecules • 4. Describe some diseases due to defects in cytoskeleton
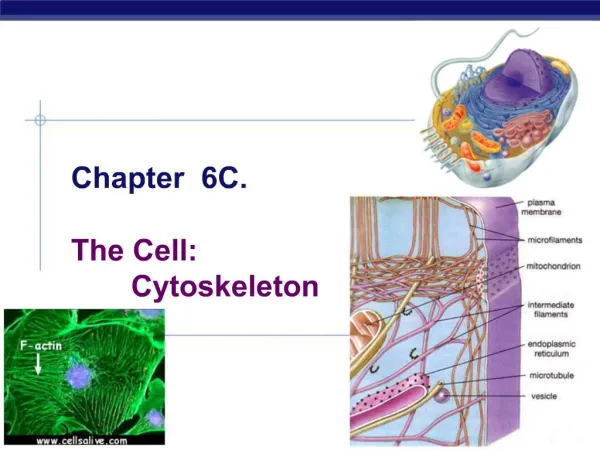
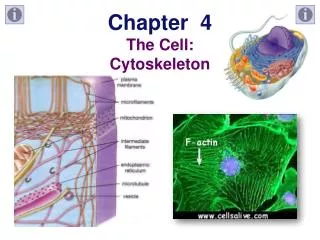
Introduction Cytoskeleton of eukaryotic cells Network of protein filaments throughout cytoplasm. Structural framework for cell shape, positions of organelles, organization of cytoplasm. Dynamic structure, continually reorganized as cells move and change shape Movement of cells, internal transport of organelles Fig. 4.31 Immunofluorescence to detect actin (blue), tubulin (yellow)
Structure and Organization of Actin Filaments Actin filaments (microfilaments – 7 nm diameter) Polymerize to actin filaments (flexible fibers, up to several µm in length) Organized into bundles, 3-D networks Actin is 375 amino acids (43 kd), highly conserved protein Abundant (5-10% cell protein) Mammals have 6 actin genes: 4 expressed in muscle cells, 2 in non-muscle cells Prokaryotic ancestor: MreB, structure for rod-shaped bacteria Fig. 12.1 Actin
Structure and Organization of Actin Filaments Actin monomer (globular [G] actin) - tight binding sites mediate head-to-tail interactions with 2 other monomers, form filaments (filamentous [F] actin). • Polarity of filaments: • All monomers oriented in same direction • Important in assembly, • In direction of myosin movement relative to actin. Fig. 12.2
Reversible Polymerization of Actin Filaments Actin reversible polymerization: Nucleation: first step of polymerization: trimer is formed, monomers added to either end. Reversible polymerization Rate monomers are added is proportional to concentration. Polymerization requires ATP, but not hydrolysis of ATP Fig. 12.3
Structure and Organization of Actin Filaments Treadmilling: Barbed end of filament grows faster Actin-ATP associates with barbed ends; ATP later hydrolyzed ADP-actin dissociates from pointed end Treadmilling at intermediate concentrations of monomers Cytochalasinsbind to barbed ends, block elongation: inhibit cell division. Phalloidin binds to filaments, prevents dissociation. (label with fluorescent dye) Fig. 12.4 Actin
Structure and Organization of Actin Filaments Actin-binding proteins regulate assembly, disassembly diverse group of proteins act in diverse ways Have ABD domains Activities of these proteins controlled by cell signals (Chapt. 15) → remodel cytoskeleton Fig. 12.5 Actin
Figs 12.6,7 Initiation of actin filaments, branches • Formin nucleates filaments to start chains, long unbranched • Formin tracks at barbed end • Tropomyosin stabilizes long filaments; • Movement requires filaments actively turn over, branch • Arp2/3 complex nucleates branches near barbed end Figs. 12.6,7 Actin
Fig 12.8 Effects of ADF/cofilin and profilin on actin filaments ADF/cofilin (actin depolymerizing factor) proteins modify existing filaments: • enhance dissociation of actin/ADP monomers from pointed end, (remain bound to monomers, prevent reincorporation). • ADF/cofilin can also sever actin filaments Profilin stimulates exchange of bound ADP for ATP so monomers available for reassembly Fig. 12.8 Actin
Fig 12.9 Actin bundles and networks Actin bundles— filaments cross-linked in closely packed parallel arrays. Actin networks—filaments cross-linked in 3-D meshwork arrays (semisolid gels) • Actin-bundling proteins are rigid (68-102 kd), cross-link, have 2 ABD • Network proteins are large (280 kd) flexible, have 2 ABD Fig. 12.9 Actin bundles, networks A is macrophage surface
Structure and Organization of Actin Bundles Parallel bundles —same polarity barbed ends at plasma membrane 14 nm apart Fimbrin in intestinal microvilli Contractile bundles — more widely-spaced (40 nm) α-actinin(102 kd dimer)cross-links motor protein myosin binds Networks: protein filamin (280 kd) flexible cross-links Filamin dimer V-shaped molecule, Actin-binding domains (ABD) each arm. Figs. 12.10,11 Actin bundles, networks
Structure and Organization of Actin Filaments Actin filaments associate with plasma membrane: 3-D network. Network and associated proteins (cell cortex) determine cell shape, involved in movement. Red blood cells (erythrocytes) model system of cortical cytoskeleton no nucleus or organelles, easy to isolate plasma membranes, associated proteins lack other cytoskeletal components Fig. 12.12
Structure and Organization of Actin Filaments Red blood cell: Spectrin: major structural protein member of calponin family of actin-binding proteins (other members include a-actinin, filamin, fimbrin, dystrophin) tetramer of two polypeptides, αand β(220, 240 kd); ends associate with short actin filaments. Fig. 12.13
Fig 12.14 erythrocyte cortical cytoskeleton binds to plasma membrane • Spectrin binds short actin chains • Ankyrin links spectrin-actin network to plasma membrane by binding to spectrin and transmembrane protein (band 3). • Protein 4.1 binds spectrin-actin junctions to transmembrane protein glycophorin. Fig. 12.14* red blood cell membrane
Structure and Organization of Actin Filaments Cytoskeleton linking proteins in other cells: Dystrophin, (427 kD), a calponin, links actin filaments to transmembrane proteins of muscle cell membranes. Transmembrane proteins link to extracellular matrix, to maintain cell stability during muscle contraction. Muscular dystrophy, X-linked inherited disease, results in progressive degeneration of skeletal muscle. Dystrophin absent or abnormal in patients with Duchenne’s or Becker’s muscular dystrophy
Structure and Organization of Actin Filaments Actin bundles attach to plasma membrane Specialized regions of plasma membrane: Contact adjacent cells, extracellular matrix, other substrata (surface of culture dish). Focal adhesions: cells attach to culture dishes: Actin bundles (stress fibers) cross-linked by a-actinin TransmembraneIntegrins Extracellular matrix proteins Tropomyosinstabilizes Talin, vinculinlink Fig. 12.15,16 actin bundles fibroblasts in culture; focal adhesions
Fig 12.17 Attachment of actin filaments to adherens junctions Adherens junctionsattach epithelial cell-cell • Continuous beltlike structure (adhesion belt) around each cell • Transmembrane proteins cadherins bind to cytoplasmic catenins, anchor actin filaments to plasma membrane • (b-catenin also signaling molecule transcriptional activator) Fig. 12.17
Figs. 12.18, 19 microvilli Actin filaments support protrusions from cell surface • Microvilli on cells for absorption Ex. microvilli of epithelial cells of intestine form layer on apical surface (brush border) ~1000 microvilli per cell; increases surface area Each microvillus: • Closely packed parallel bundles 20 to 30 actin filaments. • Filaments cross-linked by fimbrin, villlin. • Actin bundles attach to plasma membrane by calcium-binding protein calmodulin in association with myosin I. Figs. 12.18,19
Fig 12.20 cell surface projections for phagocytosis and movement Pseudopodia: phagocytosis, movement of amoebae Lamellipodia broad, sheetlike extensions at the leading edge of fibroblasts. Microspikes or filopodia, thin projections from cell Fig. 12.20: A, macrophage and tumor cell; B, amoeba, C, tissue culture cell
Fig 12.21 Structure of muscle cells 2. Myosin and cell movement: • Myosin - prototype molecular motor —converts chemical energy (ATP) to mechanical energy • Muscle contraction model: actin-myosin interactions and the motor activity of myosin molecules • Muscle fibers (50 um diam) • Fused muscle cells • Myofibrils (cytoplasm) • Thick myosin filaments • Thin actin filaments • Sarcomeres • Individual contractile units Fig. 12.21: Muscle cell
Fig 12.22 Structure of sarcomere Fig. 12.22 EM of muscle Sarcomere structure • Dark bands reflect presence or absence of myosin • Actinfilaments attached at barbed ends to Z disc, includes cross-linking protein α-actinin • Titin (extremely large, 3000 kd); single titin molecules extend from M line to Z disc; act like springs on myosins • Nebulin filaments associate with actin; regulate assembly of actin – act as rulers of length Fig. 12.23
Fig 12.24 Sliding filament model of muscle contraction Sliding filament model of muscle contraction • During contraction, sarcomere shortens, bringing Z discs closer together. • No change in width of A band;Actin moves into A band (H zone) Molecular basis: reversible binding of myosin to actin filaments: myosin motor drives filament sliding. Fig. 12.24
Fig 12.26 Organization of myosin thick filaments Muscle Myosin (Myosin II): • 500 kd (4500 aa) • 2 heavy chains – coil • 2 pairs of light chains: • regulatory, essential Thick muscle filaments: • Several hundred myosins, parallel staggered array. • Globular heads bind actin, form cross-bridges between thick and thin filaments. • Orientation of filaments reverses at M-line. Figs. 12.25,26
Fig 12.27 Model for myosin action Model of myosin action: • Reversible conformation of myosin – binds actin, ATP • ATP hydrolysis powers dissociation of actin-myosin complex • Sliding Fig. 12.27
Fig 12.28 Association of tropomyosin, troponins with actin Skeletal muscle contractiontriggered by nerve impulses: • Release of Ca2+ from sarcoplasmic reticulum (ER) • Increased [Ca2+] in cytosol affects actin binding proteins: • tropomyosin and troponin complex • Binding of Ca2+ to troponin C shifts complex, allows contraction by exposing myosin binding sites Fig. 12.28
Fig 12.29 Contractile assemblies in nonmuscle cells Nonmuscle cells have similar contractile assemblies: • Actin and Myosin II • Contraction by sliding actin filaments relative to one another. • Ex, stress fibers and adhesion belts (Figs. 12.16, 17) • Cytokinesis after mitosis (Fig. 12.30) • Membrane-bound myosin under membrane Figs. 12.29,30
Fig 12.31 Regulation of myosin by phosphorylation Contraction in nonmuscle cells, smooth muscle: • Regulated by phosphorylation of a myosin light chains. • Catalyzed by myosin lightchain kinase (MLCK) • Regulated by Ca2+-binding protein calmodulin. Fig. 12.31
Other Non-muscle myosins Oother non-muscle myosins: Myosin I - smaller than myosin II (110 kd); globular head acts as molecular motor; short tails bind other structures • Movement of myosin I along actin filament transports cargo, such as vesicle 12 other nonmusclemyosins (III - XIV) Myosin V is two-headed myosin • transports organelles, other cargo (intermediate filaments) toward barbed ends. Fig. 12.32,33
Fig 12.34 Cell migration Cell locomotion: • Extensions of plasma membrane driven by dynamic properties of actin cytoskeleton • Amoeba, • Migration of embryonic cells, • White blood cells into tissues • Cells for wound healing Inhibition of actin polymerization blocks formation of cell surface protrusions. Fig. 12.34
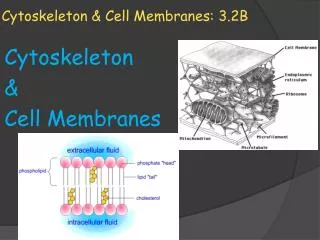
Intermediate Filaments 3. Intermediate filaments - 8-11 nm diameter • Not directly involved in cell movements • Mechanical strength, scaffold for localization of cell processes • Nuclear lamina
Intermediate Filaments Common structure: assembly Figs. 12.36, 37 See also Fig. 9.4
Intermediate Filaments Intermediate filaments Not distinct ends More stable, not dynamic behavior of actin filaments Phosphorylation can regulate assembly and disassembly (ex. nuclear lamins disassemble in mitosis) Network in cytoplasm Associate with other elements of cytoskeleton → scaffold Fig. 12.38; Network of keratins (Ab)
Intermediate Filaments Intermediate filaments function in contacts, such as epithelial cells Desmosomes— junctions to adjacent cells. Keratin filaments attach to dense protein plaques on cytoplasmic side. Attachments mediated by plakins; Transmembranecadherins link cells Hemi-desmosomes— junctions to underlying connective tissue (Fig. 12.39) Keratin filaments attached to different plakins (e.g. plectin) Transmembraneintegrins link to extracellular matrix. Fig. 12.38; Desmosome
Fig 12.40 EM of plectin bridges between intermediate filaments, microtubules • Plakins link intermediate filaments to other cytoskeleton elements • Ex. Plectin binds actin filaments, microtubules, forms bridges between them and intermediate filaments. • Increases mechanical stability of cell. Fig. 12.40 Plectin, green; Ab yellow, IF blue, microtubules, red
Fig 12.41 Experimental demonstration of keratin function Transgenic mice with mutant keratin 14: evidence for importance of intermediate filaments • Truncated keratin disrupted formation of normal keratin cytoskeleton → severe skin abnormalities. Fig. 12.41; Normal skin (top) TG skin (lower)
Intermediate Filaments & disease Human diseases - disorders of intermediate filaments: Epidermolysisbullosa simplex (EBS) patients develop skin blisters from cell lysis after minor trauma; keratin gene mutations Amyotrophic lateral sclerosis (ALS) - progressive loss of motor neurons, muscle atrophy and paralysis. Abnormalities of neurofilaments (NF-L, NF-H) Hutchinson-Gilford (Progeria) causes premature aging, involves mutations affect Lamin A protein (Chapt. 9)
Fig 12.42 Structure of microtubules 4. Microtubules - rigid hollow rods 25 nm • Dynamic structures undergo continual assembly, disassembly: cell movements, cell shape. • Globular protein (55 kd) tubulin • Tubulin dimers of α-tubulin and β-tubulin encoded by related genes • 13 protofilaments • hollow core • 25 nm diameter • Polarity: - end, + end • GTP Fig. 12.42
Fig 12.43 Treadmilling, role of GTP in microtubules Treadmilling of Microtubules: • Tubulin dimers with GTP bound to β-tubulin associate with the growing end. • After polymerization, GTP hydrolyzed to GDP, makes tubulin less stable; dimers at minus end disassociate. Fig. 12.43
Fig 12.44 Dynamic instability of microtubules Rapid GTP hydrolysis results in dynamic instability: • High concentrations tubulin-GTP: dimers added more rapidly than GTP is hydrolyzed, and microtubule grows. • Low concentration of tubulin-GTP: GTP at plus end is hydrolyzed, dimers are lost. Fig. 12.44 - End + end
Fig 12.45 Intracellular organization of microtubules Most microtubules extend from centrosome (animals) Need rapid remodeling, as for mitosis and spindle formation: Drugs colchicine and colcemid affect microtubule assembly; experimental tools, cancer treatments Vincristine and vinblastine inhibit microtubule polymerization, inhibit rapidly dividing cells, cancer chemotherapy Taxol stabilizes microtubules, blocks cell division. [plant cells do not have centrosome; microtubules extend from nucleus] Fig. 12.45
Fig 12.46 Growth of microtubules from centrosome Centrosome is microtubule-organizing center: • Minus ends of microtubules anchored: colcemid disassemble microtubules • after drug is removed, microtubules grow • - Key protein is special γ-tubulin, initiates microtubules formation Pairedcentrioles: perpendicular; amorphous pericentriolar material. • Centrioles are cylindrical; • 9 triplets of microtubules; • Lots of other proteins • Centrioles also in basal bodies of cilia, flagella Fig. 12.46 Ab to tubulin; Fig. 12.47,48 centrioles
Fig 12.49 Stability of microtubules in nerve cells Stability of microtubules: • Modulated by modifications of tubulin, e.g. phosphorylation • Microtubule-associated proteins interact: • Stabilize by capping ends of microtubules • Disassemble by sever microtubules, depolymerize • Track +-end (bind tubulin/GTP) and focus cell location • MAPs are also regulated by phosphorylation: Ex. Nerve cells: (axons, dendrites): • microtubules organized differently • distinct MAPs each type: • MAP-1, -2, tau; MAP-4 Fig. 12.49
Microtubule Motors and Movement 5. Microtubule motors and movement: 2 families of motor proteins (kinesinsand dyneins)power movements involving microtubules: Position vesicles, organelles, beating cilia, flagella, mitosis Kinesin 1: 380 kd (2 heavy chains, 2 light chains) head binds ATP, microtubule; tail binds vesicles; move to + end Dynein: 2000 kd (2-3 heavy chains, other chains) head binds ATP, microtubule; tail binds organelles; move to – end Both use ATP energy Fig. 12.50
Microtubule Motors and Movement Video-enhanced microscopy to study movements: Especially vesicles, organelles along giant squid axon Nerve cell axons may extend more than meter in length; Ribosomes only in cell body and dendrites, so proteins, membrane vesicles, etc. transported from cell body to axon Kinesin carries secretory vesicles with neurotransmitters from Golgi to terminal branches of axon. Cytoplasmic dynein transports endocytic vesicles back to cell body Fig. 12.49
Fig 12.51 Transport of vesicles along microtubules • Microtubules usually oriented with - end in centrosome; + end extends toward periphery. • Members of kinesin and dynein families transport cargo in opposite directions • Microtubules, associated motor proteins position organelles in cell. • Ex.: ER extends to periphery of cell in association with microtubules, which involves kinesin I. • Drugs that depolymerize microtubules cause ER to retract toward cell center. Figs. 12.51, 52
Microtubule Motors and Movement Cilia and flagella microtubule-based projections of plasma membrane responsible for movement of many eukaryotic cells. Similar structure: Cilia beat back-and-forth. Flagella longer, wavelike pattern of beating [Some bacteria have flagella - very different structure: protein filaments projecting from cell surface] Figs. 12.51, 52 Paramecium, Cilia, sperm
Fig 12.54 Structure of axoneme of cilia and flagella Axoneme of cilia and flagella: 250 nm diameter • Microtubules in “9 + 2” pattern: central pair, 9 outer doublets. • Complete A and partial B (10–11 protofilaments) fused to A • Nexin links microtubules; 2 arms of dynein to each A tubule Movement of cilia and flagella • Sliding outer microtubule doublets relative to one another • Powered by motor activity of axonemaldyneins. • Dynein bases bind A tubules, • Head groups bind adjacent B tubules Fig. 12.54, 56
Fig 12.55 Electron micrographs of basal bodies Minus ends of microtubules anchored in basal body, • Similar structure to centriole: 9 triplets of microtubules. • Basal bodies initiate growth of axonemal microtubules, • Anchor cilia and flagella to surface of cell. Fig. 12.55
Microtubule Motors and Movement Mitosis reorganizes microtubules Interphase microtubules disassemble free tubulin subunits reassembled to form mitotic spindle Centrosome duplicate to form 2 microtubule-organizing centers (poles) 4 types microtubules in spindle: Kinetochore Chromosomal Polar Astral Fig. 12.57, 58
Fig 12.59 Anaphase A chromosome movement Chromosome movement in Anaphase: Anaphase A — chromosomes move toward poles along kinetochore microtubules, which shorten Motor proteins Directed to Minus end help Kinesins help depolymerize microtubules Fig. 12.59