Download
1 / 66
700 likes | 1.32k Vues
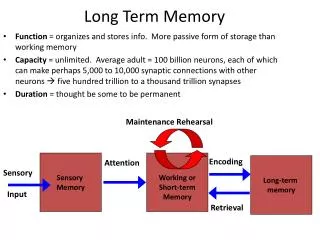
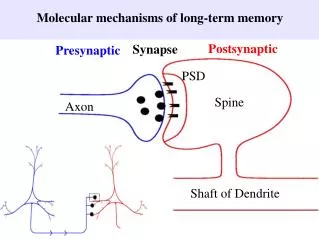
Postsynaptic. Synapse. Presynaptic. PSD. Spine. Axon. Shaft of Dendrite. Molecular mechanisms of long-term memory . LTP protocol induces postynaptic influx of Ca 2+. Postsynaptic current. Time (mins). 60. 0. LTP: an increase in synaptic strength. Long-term potentiation (LTP).

E N D
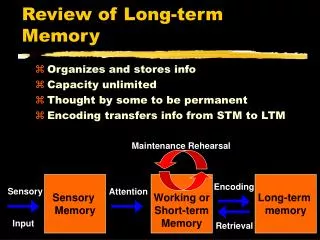
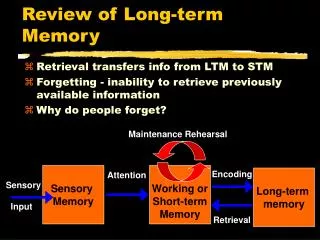
Postsynaptic Synapse Presynaptic PSD Spine Axon Shaft of Dendrite Molecular mechanisms of long-term memory
LTP protocol induces postynaptic influx of Ca2+ Postsynaptic current Time (mins) 60 0 LTP: an increase in synaptic strength Long-term potentiation (LTP) Bliss and Lomo J Physiol, 1973
LTP: an increase in synaptic strength Long-term potentiation (LTP) LTP protocol induces postynaptic influx of Ca2+ with CaMKII inhibitor or knockout Postsynaptic current Time (mins) 60 0 Lledo et al PNAS 1995, Giese et al Science 1998
Calcium-calmodulin dependent kinase II (CaMKII) One holoenzyme = 12 subunits Kolodziej et al. J Biol Chem 2000
Model of bistability in the CaMKII-PP1 system: autocatalytic activation and saturating inactivation. a) Autophosphorylation of CaMKII (2 rings per holoenzyme): P0 P1 slow P2 P1 fast Lisman and Zhabotinsky, Neuron 2001
b) Dephosphorylation of CaMKII by PP1 (saturating inactivation) = phosphatase, PP1 E k1 k2 k-1 Total rate of dephosphorylation can never exceed k2.[PP1] Leads to cooperativity as rate per subunit goes down Stability in spite of turnover
Bistability in total phosphorylation of CaMKII [Ca2+]=0.1M (basal level) Rate of dephosphoryation Rate of phosphorylation Total reaction rate 0 0 No. of active subunits 12N
Phosphorylation dominates at high calcium [Ca2+] = 2M (for LTP) Rate of dephosphoryation Rate of phosphorylation Total reaction rate 0 0 No. of active subunits 12N
The “Normal” State of Affairs (one stable state, no bistability)
How to get bistability 1) Autocatalysis: k+ increases with [C] 2) Saturation: total rate down, (k-)[C], is limited
Reaction pathways 14 configurations of phosphorylated subunits per ring P0 P1 P2 P3 P4 P5 P6
Phosphorylation to clockwise neighbors P0 P1 P2 P3 P4 P5 P6
Phosphorylation to clockwise neighbors P0 P1 P2 P3 P4 P5 P6
Random dephosphorylation by PP1 P0 P1 P2 P3 P4 P5 P6
Random dephosphorylation by PP1 P0 P1 P2 P3 P4 P5 P6
Random turnover included P0 P1 P2 P3 P4 P5 P6
Stability of DOWN state = PP1 enzyme
Stability of DOWN state = PP1 enzyme
Stability of DOWN state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Protein turnover = PP1 enzyme
Stability of UP state with turnover = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Stability of UP state = PP1 enzyme
Small numbers of CaMKII holoenzymes in PSD Petersen et al. J Neurosci 2003
Simulation methods Stochastic implementation of reactions, of rates Ri(t) using small numbers of molecules via Gillespie's algorithm: 1) Variable time-steps, ∆t: P(∆t) = ∑Ri exp(-∆t ∑Ri) 2) Probability of specific reaction: P(Ri) = Ri/∑Ri 3) Update numbers of molecules according to reaction chosen 4) Update reaction rates using new concentrations 5) Repeat step 1)
System of 20 holoenzymes undergoes stable LTP 1 Pulse of high Ca2+ here Fraction of subunits phosphorylated 0 0 20 10 Time (yrs)
Slow transient dynamics revealed Fraction of subunits phosphorylated Time (mins)
Spontaneous transitions in system with 16 holoenzymes Fraction of subunits phosphorylated Time (yrs)
Spontaneous transitions in system with 4 holoenzymes Fraction of subunits phosphorylated Time (days)
Average lifetime between transitions increases exponentially with system size
Large-N limit, like hopping over a potential barrier Reaction rates Effective potential 12N No. of active subunits 0
1) Chemical reactions in biology: x-axis = “reaction coordinate” = amount of protein phosphorylation 2) Networks of neurons that “fire” action potentials: x-axis = average firing rate of a group of neurons
Why is this important? Transition between states = loss of memory Transition times determine memory decay times.
Something like physics Barrier height depends on area between “rate on” and “rate off” curves, which scales with system size.
Physics analogy: barriers with noise ... Inherent noise because reactions take place one molecule at a time. Rate of transition over barrier decreases exponentially with barrier height ... (like thermal physics, with a potential barrier, U and thermal noise energy proportional to kT ) ?
General result for memory systems Time between transitions increases exponentially with scale of the system. Scale = number of molecules in a biochemical system = number of neurons in a network Rolling dice analogy: number of rolls needed, each with with probability, p to get N rolls in row, probability is pN time to wait increases as (1/p)N = exp[N.ln(1/p)]
Change of concentration ratios affects balance between UP and DOWN states. System of 8 CaMKII holoenzymes: 9 PP1 enzymes 7 PP1 enzymes Phosphorylation fraction Time (yrs) Time (yrs)