Download

1 / 44
440 likes | 463 Vues
Delve into the chemical principles of protein structure, amino acid classifications, functional group pKa values, stereochemistry, and unique modifications. Learn about peptide bonding and the primary structure of proteins.

E N D
1 Core 6100 Chemical Principles of Biological Systems Jim Maher GU 1611c maher@mayo.edu 4-9041 Lecture 1:Chemical principles of protein structure
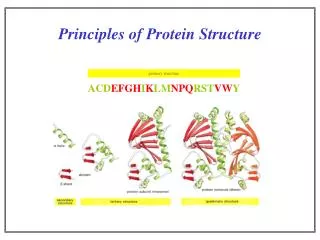
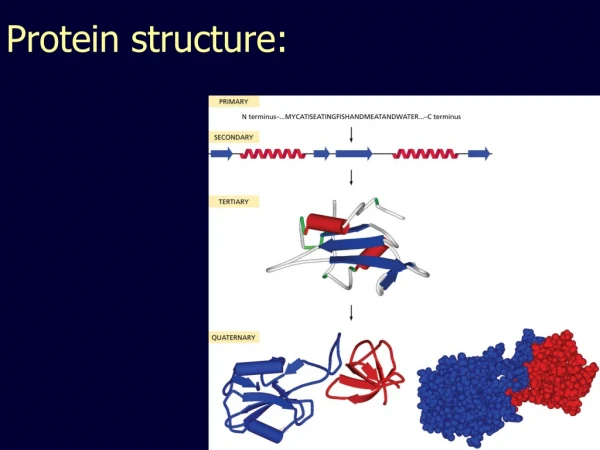
2 outline Amino acid and peptide chemistry and stereochemistry Protein primary structure (sequence) Protein secondary structure (local folding) Protein tertiary and quaternary structure (global folding) Protein spectroscopic properties reading: Lehninger 4th edition (2004): Part I: amino acids and proteins
3 Prerequisites Memorize the structures and chemical properties of the 20 natural amino acids. Memorize the triple and single letter amino acid abbreviations. Review the concept of Lewis structures (general chemistry) Review the concept of chirality (general chemistry)
S O N H C S O N C H - + - N S O H+ H S H O
4 1. Amino acid and peptide chemistry There are 20 common amino acids that appear in proteins. There are also many modified amino acids that occur in specialized proteins. Examples include 4- and 5-hydroxy proline (collagen), -carboxyglutamate (“gla domains” in blood coagulation proteins), trimethyllysine (calmodulin, histones, transcription factors), N-acetyllysine (histones), phosphoserine, phosphothreonine, phosphotyrosine, phosphohistidine, phospholysine (signal transduction) What are the functions of these weird modifications? How do they get there? These are some of the questions we’ll ask after our introductory lectures on proteins.
5 Amino acid chemical classification As you know well, amino acids can be classified according to the chemical properties of their side chains: Nonpolar Polar-charged (acidic vs. basic) Polar-uncharged How strongly do amino acid functional groups bind protons? pKa value of a functional group tells us the pH where that group is 50% protonated. High pKa = tight proton binding (bases). Low pKa = weak proton binding (acids).
6 pKa values for amino acid functional groups
7 pKa values for amino acid functional groups Acetic acid (pKa 4.76) can be considered a model for the carboxyl group. Methylammonium ion (pKa 10.6) can be considered as a model for the amino group. Therefore, amino acids are zwitterions at pH 7 Interestingly, the pKa values of the amino and carboxyl groups of amino acids are lower (protons easier to remove) than the model compounds. pKa2 9.78 (not 10.6) pKa1 2.35 (not 4.76) H+ H+ +1 0 -1
8 pKa values for amino acid functional groups Acetic acid (pKa 4.76) can be considered a model for the carboxyl group. Methylammonium ion (pKa 10.6) can be considered as a model for the amino group. Therefore, amino acids are zwitterions at pH 7 Interestingly, the pKa values of the amino and carboxyl groups of amino acids are lower (protons easier to remove) than the model compounds. pKa2 9.78 (not 10.6) pKa1 2.35 (not 4.76) H+ H+ +1 0 -1
9 pKa values for amino acid functional groups pKa2 9.78 (not 10.6) pKa1 2.35 (not 4.76) H+ H+ +1 0 -1 pKa1 lower due to inductive effect (anionic carboxylate stabilized by cationic ammonium). It thus seems strange that pKa2 is lower if inductive effects dominate. However, when pH is high near pKa2, the cost of solvation of the zwitterion is higher, slightly favoring the anion. Water dipoles
9 pKa values for amino acid functional groups pKa2 9.78 (not 10.6) pKa1 2.35 (not 4.76) H+ H+ +1 0 -1 pKa1 lower due to inductive effect (anionic carboxylate stabilized by cationic ammonium). It thus seems strange that pKa2 is lower if inductive effects dominate. However, when pH is high near pKa2, the cost of solvation of the zwitterion is higher, slightly favoring the anion. repulsion Water dipoles repulsion
10 Amino acid stereochemistry R/S system: Number from high -> low. Rotate so lowest faces away. Determine relationship 1-3 2 1 S 3 4 CCW: S CW: R Glycine: not chiral. Cysteine: R. All others: S. D/L system: All amino acids except glycine are called “L” by tradition and analogy with the reference compound, L-glyceraldehyde, where solutions rotate plane-polarized light in left counter-clockwise (left) direction. In fact, not all chiral amino acid solutions do this.
11 Amino acid stereochemistry All amino acids that appear in proteins by ribosomal synthesis have the “L” configuration. A wide variety of biological peptides contain D-amino acids, but these are created by non-ribosomal synthases. Examples: gramicidin, cyclosporine, valinomycin. Cyclosporine (immunosuppressant)
12 Proteins are not always composed only of amino acids!
13 2. Primary structure: peptide and amino acid covalent bonding Polypeptide chains include several kinds of important covalent bonds between amino acids. Peptide (amide) bond: amounts to a condensation reaction between carboxyl and amino groups of two amino acids. Actually catalyzed by ribosome using tRNA esters. -H2O + +H2O peptide (=amide) bond Arrow lengths in equilibrium reflect reaction thermodynamics at pH 7
14 2. Primary structure: peptide and amino acid covalent bonding Disulfide (cystine) bond: Formed by 2 e- oxidation of two cysteine residues -2e- disulfide bond -2H+
15 2. Primary structure: Condensing several amino acids into a linear polymer creates a polypeptide. Draw by starting with a skeleton and then decorating.
15 2. Primary structure: Condensing several amino acids into a linear polymer creates a polypeptide. Draw by starting with a skeleton and then decorating. chiral centers remain “L”
15 2. Primary structure: Condensing several amino acids into a linear polymer creates a polypeptide. Draw by starting with a skeleton and then decorating. N-terminus C-terminus chiral centers remain “L” Recognize the “N” and “C” termini
16 2. Primary structure: Key Point: although drawn as a single bond, the bond between the carbonyl carbon and amide nitrogen has partial double-bond character. Consider the resonance structures: The structure on the right includes a C-N double bond. These Lewis resonance structures convey the idea that the actual peptide bond structure amounts to a hybrid of these resonance structures, weighted by importance. The amide bond has partial (~40%) double-bond character.
17 2. Primary structure: The double-bond character of the amide bond can also be seen from model compounds: methylamine formamide 1.474 Å 1.376 Å The C-N bond in formamide is significantly shorter than in methylamine. Similar trends are observed in infrared stretching frequencies:
18 2. Primary structure: The double-bond character of the amide (peptide) bond is of crucial importance in understanding polypeptide folding. There is a strong barrier to rotation around the C-N bond. Amides therefore are relatively rigid and exist in two distinct conformations. trans (more stable) cis (less stable) Due to steric interactions between side chains, the cis conformation is somewhat less favored (by ~ 2 kcal/mol) and is therefore less frequent in folded polypeptides.
19 2. Primary structure: Because of the rigidity of amide bonds, polypeptides conformations can be viewed by focus on the alpha carbon atoms, with each C-C(O)-N unit treated as a plane.
19 2. Primary structure: Because of the rigidity of amide bonds, polypeptides conformations can be viewed by focus on the alpha carbon atoms, with each C-C(O)-N unit treated as a plane.
19 3. Primary structure: Because of the rigidity of amide bonds, polypeptides conformations can be viewed by focus on the alpha carbon atoms, with each C-C(O)-N unit treated as a plane. Two such flanking planes can have many relative rotary orientations, defined by angles (phi: upstream) and (psi: downstream). N C C(O) Both angles are defined as 180˚ for an all-trans chain.
+180 0 -180 0 +180 19b 2. Primary structure: Because of steric side-chain collisions, only certain families of conformations (pairs of and ) are allowed for real proteins. This can be represented by a Ramachandran plot: Limited options for combinations of and Wikipedia data for PCNA
+180 0 -180 0 +180 19b 2. Primary structure: Because of steric side-chain collisions, only certain families of conformations (pairs of and ) are allowed for real proteins. This can be represented by a Ramachandran plot: glycines Wikipedia data for PCNA http://www.biochem.ucl.ac.uk/~roman/procheck/manual/examples/plot_02a.gif
Lecture 1 Review 1˚ 2˚ 3˚ 4˚ amino acids and ionization (pKa values) peptide (amide) bonds -partial double-bond character (why?) -not free to rotate -phi () and psi () angles -Ramachandran plots 2˚ (secondary) structures: stabilized by H-bonds -helix: right-handed spiral 3.6 aa/turn dipole (+ at amino end of helix) favored for some aa, broken by others -sheet: silk example parallel, antiparallel R groups out of plane collagen or “L” triple helix: left handed
Lecture 1 Review 1˚ 2˚ 3˚ 4˚ 3˚ (tertiary) structures: packing of 2˚ structure units -challenge of prediction -in vitro vs. in vivo ? 4˚ (quaternary) structure: non-covalent -self-assembly of multiple subunits
A D H 20 3. Secondary structure: 1˚ structure: linear amino acid sequence, N -> C 2˚ structure: local regular structures. In proteins these structures are stabilized by backbone Hydrogen bonds not involving side chains. Hydrogen bonding (H-bonding) involves favorable non-covalent interactions between an H-bond donor hydrogen, and (typically) a heteroatom with at least one lone pair of valence electrons not involved in a covalent bond. In polypeptides, 2˚ structure involves backbone H-bonds between a lone pair of electrons on the carbonyl oxygen and the hydrogen atom of the amide nitrogen
21 3. Secondary structure: -Helix. A familiar 2˚ structure element observed when and are in the range -50˚ to -60˚. The backbone spirals in a right-handed fashion (N -> C) along an axis defined by the polypeptide, while the side chains project down and out from the axis. The -helix has about 3.6 residues per turn, so every 7th residue is roughly on the same helical face. Let’s look at some examples from -helices in the dimeric bZIP yeast DNA binding protein GCN4: Note: what is the plural form of the word “helix” ?
1 5 4 2 0 6 7 3 C 22 N PDB: 1ysa
23 3. Secondary structure: -Helix. Since all H-bonds in an -helix have dipolar character and align in the same direction along the helix axis, the overall -helix has an associated dipole representing the vector sum of the individual dipoles of each peptide. This dipole amounts to an additional partial positive charge at the N-terminal end of the -helix, and an additional partial negative charge at the C-terminal end. C -0.42 +0.42 Net charge = 0 But dipole moment: -0.2 +0.2 N
24 3. Secondary structure: Certain amino acids (A, L, M, H, Q, V) are preferred in -helices, while others (P, G, Y, S) tend to disrupt. In particular, the pyrrolidine ring of proline forms a rigid structure that bends the polypeptide backbone and tends to disrupt -helices.
25 3. Secondary structure: An interesting example of conditional-helical 2˚ structure is observed for poly-L-lysine or poly-L-glutamic acid. These form stable -helices (as detected by circular dichroism spectroscopy: CD) only when their side chains are uncharged. Why? Redrawn from P. Doty in J.L. Oncley (ed.) Biophysical Science, p. 108, John Wiley & Sons, New York, 1959
26 3. Secondary structure: -sheet. A second familiar 2˚ structure element is observed when and are in the range of 180˚. -sheets are made up of parallel or antiparallel extended polypeptide strands sharing backbone H-bonds between strands. antiparallel parallel N C C N C C N N Note: positions of “R” groups (side chains). Stereochemistry remains unchanged. H-bonds shown as dashes.
27 3. Secondary structure: -sheet. Examples from thioredoxin +side chains 2 backbones
28 3. Secondary structure: *right-handed triple helix of left-handed helices Other cases. Less common 2˚ structures exist, as well as loops or coils (often flexible, on protein surfaces, and less conserved) as well as turns or bends.
29 4. Tertiary (3˚) and quaternary (4˚) structure: 3˚ structure: the arrangement of 2˚ structural elements in space to yield a folded polypeptide chain. 4˚ structure: the organization of separate polypeptide chains into a protein complex with multiple subunits. These higher-order folding interactions are driven by side-chain interactions including hydrophobic, van der Waals, and electrostatic forces, as well as H-bonds and (rarely) covalent bonds. The ability to predict 2˚ - 4˚ protein structure from 1˚ sequence is a long-standing challenge in biochemistry.
30 5. Self-assembly: Among the remarkable features of proteins is the observed ability of some complex systems to self-assemble. This implies a process in which all steps have both favorable free energies and permissive kinetics. An impressive example are the dozens of proteins of bacteriophage T4. How can such a system self-assemble without energy input? NSF
31 5. Spectroscopic properties of proteins UV/visible absorbance properties. The important chromophores are the aromatic amino acids tryptophan, tyrosine, and phenylalanine. The oxidized form of cysteine (cystine) also absorbs UV light. Note: log scale Jakubowski CSBSJU Biochem.arizona.edu
32 5. Spectroscopic properties of proteins UV/visible absorbance properties. Tryptophan wins.
32 5. Spectroscopic properties of proteins If you know the amino acid composition, the protein concentration can be estimated from the UV absorbance, but the molar extinction coefficients actually vary depending on amino acid environment (pH, solvent exposure). One approach is to determine the ratio of absorbance under native conditions vs. denatured in 6 M guanidine HCl (all side chains equally exposed) for a given protein, and subsequently apply this correction when measuring samples of that native protein.