Two Substrate Reactions
Two Substrate Reactions. Many enzyme reactions involve two or more substrates. Though the Michaelis-Menten equation was derived from a single substrate to product reaction, it still can be used successfully for more complex reactions (by using k cat ). Random. Ordered. Ping-pong.


Two Substrate Reactions
E N D
Presentation Transcript
Two Substrate Reactions • Many enzyme reactions involve two or more substrates. Though the Michaelis-Menten equation was derived from a single substrate to product reaction, it still can be used successfully for more complex reactions (by using kcat). Random Ordered Ping-pong
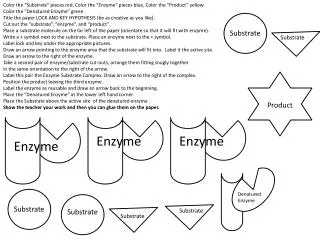
Two Substrate Reactions • In random order reactions, the two substrates do not bind to the enzyme in any given order; it does not matter which binds first or second. • In ordered reactions, the substrates bind in a defined sequence, S1 first and S2 second. • These two reactions share a common feature termed a ternary complex, formed between E, ES1, ES2 and ES1S2. In this situation, no product is formed before both substrates bind to form ES1S2.
Two Substrate Reactions (cont) • Another possibility is that no ternary complex is formed and the first substrate S1 is converted to product P1 before S2 binds. These types of reactions are termed ping-pong or double displacementreactions.
The catalytic mechanism of chymotrypsin: a member of the serine protease family; catalyzes the hydrolytic cleavage of peptide bonds adjacent to aromatic amino acid residues (with a rate enhancement of at least 109). Principles illustrated: Transition-state stabilization; General acid-base catalysis; Covalent catalysis.
Chymotrypsin(and other proteins) are activated via proteolytic cleavage of precursor proteins (zymogens or preproteins).Many proteases activated this way can be inactivated by inhibitor proteins tightly-bound in the active sites.
Active chymotrypsin and trypsin are produced from inactive zymogens via proteolytic cleavage, with conformational changes exposing the active sites.
The catalytically important groups of chymotrypsin were identified by chemical labeling studies • Organic fluorophosphates such as diisopropylphosphofluoridate (DIPF) irreversibly inactivate chymotrypsin (and other serine proteases) and reacts only with Ser195 (out of the 25 Ser residues).
A second catalytically important residue, His57, was discovered by affinity labeling with tosyl-L-phenylalanine chloromethylketone (TPCK) •TPCK alkylates His 57 • Inactivation can be inhibited by b-phenylpropionate (competitive inhibitor) • TPCK modification does not occur when chymotrypsin isdenatured in urea.
Rapid initial burst kinetics indicates an acyl-enzyme intermediate • The kinetics of chymotrypsin is worked out by using artificial substrates (esters), yielding spectroscopic signals upon cleavage to allow monitoring the rate of reactions. Colorless substrate Yellow product Fast Slow This reaction is far slower than the hydrolysis of peptides! Km = 20 mM Kcat = 77 s-1
The catalysis of chymotrypsin is biphasic as revealed by pre-steady state kinetics Slow phase (enzymes will be able to act again only after a slow deacylation step) “burst” (fast) phase (rapid acylation of all Enzymes leading to release of p-nitrophenol) Milliseconds after mixing
Determination of the crystal structure of chymotrypsin (1967) revealed a catalytic triad: Ser195, His57, Asp102.
Chymotrypsin: three polypeptide chains linked by multiple disulfide bonds; a catalytic triad. Active site His57 Ser195 Asp102 Cleft for binding extended substrates Trypsin, sharing a 40% identity with chymotrypsin, has a very similar structure.
A catalytic triad has been found in all serine proteases: the Ser is thus converted into a potent nucleophile
The Peptide Bond has partial (40%) double bond character as a result of resonance of electrons between the O and N The hydrolysis of a peptide bond at neutral pH without catalysis will take ~10-1000 years!
Chymotrypsin (and other serine proteases) acts via a mixture of covalent and general acid-base catalysis to cleave (not a direct attack of water on the peptide bond!)
Formation of the ES complex Asp102 functions only to orient His57. E S Formation of ES1 ES1 The peptide bond to be cleaved is positioned by the binding of the side chain of an adjacent hydrophobic residue in a special hydrophobic pocket.
His57acts as a general base in deprotonating Ser195, the alkoxide ion then acts as a nucleophile, attacking the carbonyl carbon. ES1 Ser195forms a covalent bond with the peptide (acylation) to be cleaved. a trigonal C is turned into a tetrahedral C. The tetrahedral oxyanion intermediate is stabilized by the NHs ofGly193andSer195 Pre-acylation Preferential binding of the transition state:oxyanion holestabilization of the negatively charged tetrahedral intermediate of the transition state. oxyanion hole
His57 acts as a general acid in cleaving the peptide bond. ES1 Acylation Releasing of P1 The amine product is then released from the active site with the formation of an acyl-enzyme covalent intermediate. Acyl-E
Entering of S2 E’S2 Acyl-E Water (the second substrate) then enters the active site.
His57acts as a general base again, allowing water to attack the acyl-enzyme intermediate, forming another tetrahedral oxyanion intermediate, again stabilized by the NHs of Gly193 and Ser195 (similar to step 2) Pre-deacylation E’S2
EP2 His57acts as a general acid again in breaking the covalent bond between the enzyme and substrate (deacylation) (similar to Step 3). Deacylation
Recovered enzyme Release of P2 E The second product (an acid) is released from the active site, with the enzyme recovered to its original state. EP2
2nd product 1st substrate E EP2 ES The proposed complete catalytic cycle of chymotrypsin (rate enhancement: 109) A Ping-Pong Mechanism 1st product E’S2 Acyl-E Deacylation phase Acylation phase 2nd substrate