Download
1 / 25
250 likes | 474 Vues
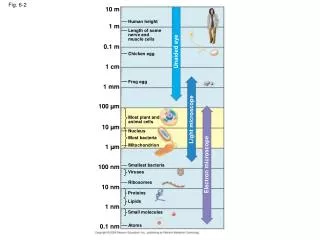
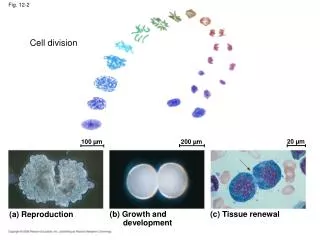
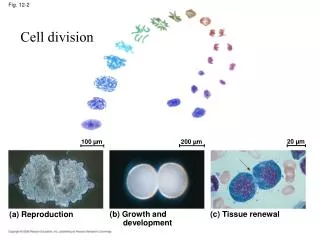
20 µm. 100 µm. 200 µm. (a) Reproduction. (b) Growth and development. (c) Tissue renewal. Cell division. Fig. 12-2. Bacterial chromosome. Prokaryotes. Chromosomes. Microtubules. Intact nuclear envelope. Dinoflagellates. Kinetochore microtubules. Intact nuclear envelope.

E N D
20 µm 100 µm 200 µm (a) Reproduction (b) Growth and development (c) Tissue renewal Cell division Fig. 12-2
Bacterial chromosome Prokaryotes Chromosomes Microtubules Intact nuclear envelope Dinoflagellates Kinetochore microtubules Intact nuclear envelope Diatoms Kinetochore microtubules Centrosome Fragments of nuclear envelope Most eukaryotes Binary fission (prokaryotes) Mitosis (eukaryotes) Types of cell divisions that produce identical offspring (clones) in various organisms LE 12-12
Cell wall Origin of replication Plasma membrane E. coli cell Bacterial chromosome Chromosome replication begins. Soon thereafter, one copy of the origin moves rapidly toward the other end of the cell. Two copies of origin LE 12-11 Origin Origin Replication continues. One copy of the origin is now at each end of the cell. Replication finishes. The plasma membrane grows inward, and new cell wall is deposited. Two daughter cells result.
0.5 µm Chromosome duplication (including DNA synthesis) Centromere Sister chromatids Separation of sister chromatids Centromeres Sister chromatids Eukaryotic genomes are larger and more complex LE 12-4
The cell cycle INTERPHASE S (DNA synthesis) LE 12-5 G1 Mitosis Cytokinesis G2 MITOTIC (M) PHASE
10 µm TELOPHASE AND CYTOKINESIS METAPHASE ANAPHASE G2 OF INTERPHASE PROPHASE PROMETAPHASE Mitosis in animal cells LE 12-6ca
Mitosis in animal cells Kinetochore Centrosomes (with centriole pairs Early mitotic spindle Chromatin (duplicated) Aster Fragments of nuclear envelope Nonkinetochore microtubules Centromere LE 12-6aa Nucleus Plasma membrane Chromosome, consisting of two sister chromatids Kinetochore microtubule Nuclear envelope
Mitosis in animal cells Nucleolus forming Metaphase plate Cleavage furrow LE 12-6ba Nuclear envelope forming Centrosome at one spindle pole Daughter chromosomes Spindle
100 µm Cleavage furrow Daughter cells Contractile ring of microfilaments Cleavage of an animal cell (SEM) Cytokinesis in animal cells LE 12-9a
Mitosis in plant cells Chromatin condensing Nucleus 10 µm Chromosomes Cell plate Nucleolus LE 12-10 Prometaphase. We now see discrete chromosomes; each consists of two identical sister chromatids. Later in prometaphase, the nuclear envelope will fragment. Prophase. The chromatin is condensing. The nucleolus is beginning to disappear. Although not yet visible in the micrograph, the mitotic spindle is starting to form. Metaphase. The spindle is complete, and the chromosomes, attached to microtubules at their kinetochores, are all at the metaphase plate. Telophase. Daughter nuclei are forming. Meanwhile, cytokinesis has started: The cell plate, which will divide the cytoplasm in two, is growing toward the perimeter of the parent cell. Anaphase. The chromatids of each chromosome have separated, and the daughter chromosomes are moving to the ends of the cell as their kinetochore micro- tubules shorten.
Cytokinesis in plant cells LE 12-9b Vesicles forming cell plate Wall of parent cell 1 µm New cell wall Cell plate Daughter cells Cell plate formation in a plant cell (TEM)
Experiment 1 Experiment 2 M S G1 G1 LE 12-13 Regulation of cell cycle M S M S When a cell in the M phase was fused with a cell in G1, the G1 cell immediately began mitosis—a spindle formed and chromatin condensed, even though the chromosome had not been duplicated. When a cell in the S phase was fused with a cell in G1, the G1 cell immediately entered the S phase—DNA was synthesized.
G1 checkpoint LE 12-14 Control system S G1 G2 M M checkpoint G2 checkpoint
LE 12-15 G0 G1 checkpoint G1 G1 If a cell does not receive a go-ahead signal at the G1 checkpoint, the cell exits the cell cycle and goes into G0, a nondividing state. If a cell receives a go-ahead signal at the G1 checkpoint, the cell continues on in the cell cycle.
S G2 M S G2 M G1 M G1 G1 Fig. 12-17a MPF activity Cyclin concentration Time (a) Fluctuation of MPF activity and cyclin concentration during the cell cycle
G1 S Fig. 12-17b Cdk Cyclin accumulation M G2 Degraded cyclin G2 checkpoint Cdk Cyclin is degraded Cyclin MPF (b) Molecular mechanisms that help regulate the cell cycle
The Fucci cell cycle visualization method. Cells in phase G1 fluoresce bright red. As they transition into the S phase and begin DNA replication, Cdt1 levels are dramatically reduced while Geminin levels increase. This results in faint yellow fluorescence early in G1/S that soon gives way to robust green fluorescence, (G2) which lasts until the cell re-enters G1 phase. Sakaue-Sawano, A.,et al. Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell132, 487–498 (2008).
Scalpels Experimental set-up for cell growth Fig. 12-18 Petri plate Without PDGF cells fail to divide With PDGF cells prolifer- ate Cultured fibroblasts 10 µm
MUTATION Growth factor Hyperactive Ras protein (product of oncogene issues signals on its own. G protein Cell cycle-stimulating pathway Receptor Protein kinases (phosphorylation cascade) NUCLEUS Transcription factor (activator) DNA Gene expression Protein that stimulates the cell cycle Signal transduction pathway for cell cycle stimulation LE 18-21
Cancer tissue lacks growth inhibition (or may have internal over stimulation (Ras mutations) Anchorage dependence Fig. 12-19 Density-dependent inhibition Density-dependent inhibition 25 µm 25 µm (a) Normal mammalian cells (b) Cancer cells
Asymmetric Inheritance of Mother Versus Daughter Centrosome in Stem Cell Division Science 26 January 2007:Vol. 315. no. 5811, pp. 469 - 470DOI: 10.1126/science.1138237
Asymmetric segregation is produced by centrosomes Fig. 1. GFP-labeled daughter centrosomes migrate away from the niche. Stereotyped positioning of centrosomes in male germline stem-cells during interphase sets up the orientation of the mitotic spindle [adapted from (6)]. Red, centrosome; blue, hub; green, tubulin. Drosophila male germline stem cells (GSCs) are maintained through attachment to somatic hub cells, which constitute the stem cell niche. Hub cells secrete the signaling ligand Upd, which activates the Janus kinase–signal transducer and activator of transcription (JAK-STAT) pathway in the neighboring germ cells to specify stem cell identity (4, 5). Drosophila male GSCs normally divide asymmetrically, producing one stem cell, which remains attached to the hub, and one gonialblast, which initiates differentiation. This stereotyped asymmetric outcome is controlled by the orientation of the mitotic spindle in GSCs: The spindle lies perpendicular to the hub so that one daughter cell inherits the attachment to the hub, whereas the other is displaced away (6).
Centrosome positions are controlled by cell junctions Fig. 3. Centrosomes next to the hub harbor robust microtubule arrays. Electron micrograph and summary diagram of a proximal centrosome in a GSC. Arrowheads in (A') show a microtubule that runs from the centrosome to the adherens junction. Science 26 January 2007:Vol. 315. no. 5811, pp. 469 - 470DOI: 10.1126/science.1138237
Centrosomin is a protein that controls centrosome segregation Fig. 4.Mutant for centrosomin (cnn) ; cnn is required for nonrandom segregation of mother and daughter centrosomes. Centrosomin (cnn) is an integral centrosomal protein required to anchor astral microtubules to centrosomes Science 26 January 2007:Vol. 315. no. 5811, pp. 469 - 470DOI: 10.1126/science.1138237
Hub cells control stem cell renewal and differentiation Model for JAK pathway activity in embryogenesis. Upd is the ligand for stimulation of the JAK pathway. Upd protein is produced in hub cells, in which it is glycosylated and secreted, and diffusion is restricted by association with the ECM. Through binding of Upd to a yet unidentified receptor, the Hop JAK is stimulated, resulting in phosphorylation of Stat92E. Ultimately, transcription of specific genes, such as eve, is activated. D. A. Harrison, P. E. McCoon, R. Binari, M. Gilman, N. Perrimon, Genes Dev.12, 3252 (1998)