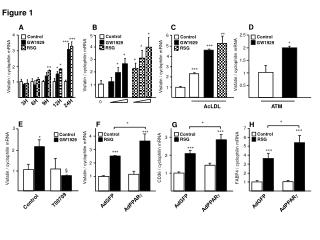
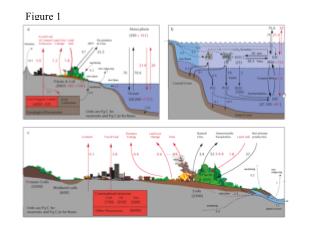
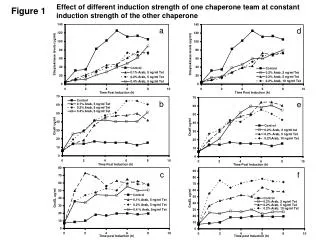
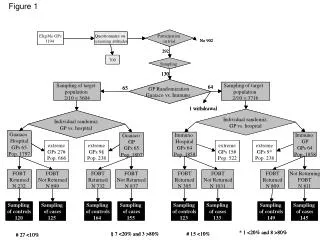
Figure 1
Class. Figure 1. Order. Figure 3. Order. Table 6 – Archaea representatives and their respective importance in each library. Figure 6. Figure 2. Figure 4. Carbon cycle. ↑ CO 2. Figure 5. ↓ Denitrifiers Streptomyces Bacillus. ↑ Respiration.


Figure 1
E N D
Presentation Transcript
Class Figure 1 Order Figure 3 Order Table 6 – Archaea representatives and their respective importance in each library Figure 6 Figure 2 Figure 4 Carbon cycle ↑ CO2 Figure 5 • ↓ Denitrifiers • Streptomyces • Bacillus ↑ Respiration Figures 5, 6 and 7 show differences in the populations of the eukaryotes isolated from the poplar rhizosphere under ambient and elevated CO2. N2 Plants ↑ Photosynthesis ≈ N fixing bacteria ↑ Plant biomass NO3 ↑ Soil oxygenation ≈ nitrate bacteria ↑ nutrient flow Figure 7 • Anaerobic populations • Archaea • Clostridia • Desulfotomaculum ↑ Fine root turnover • ≈ Nitrite bacteria • Nitrosospora NO2 • ↓ Microbial diversity • Heterobasidiomycetes • Zygomycetes • Archaea • ↓ Plant pathogens • Urediniomycetes • ↑ Decomposers • Ectomycorrhizal fungi • Actinobacteria • Sphingobacteria • Flavobacteria NH4+ Nitrogen cycle Xrepresents blockage due to increase rhizosphere oxygenation Data set Clustering method Percent Similarity Edit distance Phylotypes observed Chao1 Index Chao1 SD Simpson Index (Equitability) Ambient Prokaryotes 99% 14 882 3,989 400.7 0.39 97% 42 811 3,042 292.6 0.33 95% 70 696 2,281 217.9 0.25 Fungi 99% 18 88 345 118.8 0.41 97% 54 60 187 66.7 0.25 95% 90 35 323 311.5 0.14 Archaea 99% 10 1209 18,580 3,001.5 0.56 97% 30 564 6,592 1,392.8 0.04 95% 50 280 2,551 695.7 0.04 Elevated Prokaryotes 99% 14 839 5,888 771.9 0.24 97% 42 803 4,854 605.3 0.21 95% 70 738 3,858 468.8 0.20 Fungi 99% 18 54 142 45.3 0.18 97% 54 39 100 35.0 0.11 95% 90 31 97 43.8 0.10 Archaea 99% 10 958 18,881 3,924.3 0.11 97% 30 362 3,431 820.7 0.02 95% 50 182 1,274 374.5 0.03 N-235Population Dynamics in the Rhizosphere: How Elevated CO2 Transforms Microbial Community Composition C. Lesaulnier 1,3*, D. Papamichail 2, S. McCorkle 1, D. Zak 4, B. Ollivier 3, S. Skiena 2, S. Taghavi 1, D. van der Lelie 1 1Brookhaven National Laboratory, Upton, NY, USA. 2Stony Brook University, Stony Brook, NY, USA. 3Institut de Recherche pour le Developpement, Marseille, FRANCE. We note an absence of Urediniomycetes under elevated CO2 which is very interesting given that they are predominantly represented and comprised of plant pathogens (11). Zygomycetes - Representatives of this phylum were observed to decrease in population importance, the most significant change occurring to representatives affiliated with Mortierella. This population was observed to decrease by 75%. Ascomycete – Though the Ascomycete population importance remained stable (i.e. number of reported clones stayed the same), we notice the disappearance of the dominant species Cazia under ambient conditions and the incurring dominance of representatives belonging to Pachyphloeus under conditions of elevated CO2. Chytridiomycetes - Representatives belonging to Chytridiomycetes appear to have been unaffected by the increase in CO2. Though it is not possible at present to extrapolate potential roles for these organisms, we notice significant rearrangements in the importance of represented species. Abstract How much is enough? Scientists from all disciplines are collaborating to address the issues of anticipating the effects of anthropogenic activity and environmental change, for the purpose of predictive ecological modeling. Though no globally accepted model exists on plant responses to atmospheric carbon dioxide (CO2), little work has been done to characterize the forest flora rhizosphere to better determine their overall potential to counterbalance the effects of this greenhouse gas. Our findings show that though total soil biomass remained unchanged for prokaryotes and eukaryotes, microbial species richness decreased and noticeable shifts in dominant genera were observed. Archaea populations were noted to decrease under elevated CO2 and we hypothesize that oxidative stress due to increased fine root production and improved rhizosphere aeration inhibits their growth. This was further supported by similar observations in strictly anaerobic bacterial populations. Our data, combined with previously documented microbial activities, provides the foundation to better interpret observed changes in plant biomass, element cycling and the sustainability of terrestrial ecosystem productivity under elevated CO2. Table 1 – Richness estimations and equitability for determining population diversity Conclusion Figures 1 - 4 show differences in the populations of the prokaryotes isolated from the poplar rhizosphere under ambient and elevated CO2. Figure 8 – Model for how nutrient cycles are affected by elevated CO2. Objectives Bacteroidetes - A 30% increase in Chitinophaga representatives was observed and these Sphingobacteria have known chitinolytic properties. A 40% increase in Flavobacterium was also observed though we do not yet know why this could be. Firmicutes - Our results for Bacillus are surprising given that their metabolic capabilities are diverse and include the degradation of almost all substrates derived from plant sources. Their capacity to carry out nitrification, nitrogen fixation and denitrification has also been noted (4,5). Results for Clostridia are due to a significant decrease in representatives clustering with Desulfotomaculum species. These obligate anaerobes seem to be inhibited under elevated CO2. Obligate organisms may cease to function competitively as their environment segues to an incompatible redox state. Furthermore, they may quickly drop out of a soil community due to the effects of O2 toxicity as the environment is forced to become aerobic (6). Though this result is preliminary, its importance due to the effects of elevated CO2 has, to date, been overlooked. Proteobacteria - The importance of Proteobacteria is consistent with other soil studies whereby they have been noted to make up approximately 40% of libraries derived from soil bacterial communities (7). Any conclusions on the metabolic capabilities within this phylum are hindered by its extent of physiological diversity. Interestingly we noted no change in the populations of β-Proteobacteria involved in nitrification, predominantly among the Nitrosospira. No significant variation in community composition among the delta-Proteobacteria, which comprises 10% of the prokaryotic community, was found under elevated CO2. This is not surprising given that they have previously been reported to tolerate microaerophillic conditions (8). Verrucomicrobia - This population has been suggested to be negatively impacted by soil moisture (9) which has been reported to increase under elevated CO2 (10) and therefore it is not surprising that we note a decrease in this aerobic heterotrophic population. In an attempt to better understand the importance and impact of plant associated microbes, we aimed to provide the first large scale detailed analysis on microbial population changes due to elevated CO2 in all three domains of life present in the rhizosphere of Populus tremuloides. Methods A total of 3000 prokaryotic, 3000 archaea 16S and 2000 eukaryotic 18S ribosomal RNA gene sequences were obtained from soil extracted DNA of the poplar rhizosphere, split equally in number from under both ambient and elevated (550ppm) atmospheric CO2 concentrations. These were then taxonomically assigned using an optimized blast classifier built from a curated data set and comparisons were then made between the two populations. Archaea Community We present the first detailed analysis on how the rhizosphere microbial community of Populus tremuloides is affected by elevated CO2. Due to the observed changes in the poplar microbial community composition we postulate that that rhizosphere conditions under elevated CO2 alters the interaction between the microorganisms in its rhizosphere. The microbial community adapts so as to sustain uptake of nitrogen and other microelements in favor of the plant. This increased soil oxygenation hinders denitrification and anaerobic processes, and results in an abundance of ectomychorrizal fungi. Though this may further assist the host plant in the uptake of essential nutrients, the decrease in Desulfotomaculum brings to light the potential to significantly alter S cycling in the rhizosphere. Our results suggest that the increase in fine root biomass production and turnover under elevated CO2 favorizes the symbiotic relationship between poplar and ectomychorrizal fungi, resulting in the out-competing of saprophytic and other ectomychorrizal fungi. The effect of oxidative stress was further addressed in the archaea population where a large decrease in population importance under elevated CO2 was observed. Results Table 2 – Quantification of domain specific soil DNA concentrations under ambient and elevated CO2. Prokaryotic Community Actinobacteria - The most significant changes included their doubling, with Arthrobacter representatives increasing 15 fold (P <0.0001). These gram-positive chemoheterotrophs have been demonstrated to play a role in the degradation of many recalcitrant forms of soil carbon and involved in nitrification (1). A 50% decrease in the Streptomyces population was observed. Representatives within this group are known to have denitrifying characteristics (2). A marked increase of 30% in Sporichthya was observed, though as yet we do not know what this could be due to. Acidobacteria - Members of the phylum Acidobacteria halved in population importance. The physiological properties of these organisms are unclear, though this observation is inconsistent with recently published results under conditions of elevated CO2 (3). Eukaryotic Community References & Acknowledgements Though total reported fungal community biomass remained constant, as determined by qPCR, significant populations changes were observed. The most significant being a shift towards the Agaricales ectomycorrhizal fungi, predominantly belonging to the genus Inocybe. These are commonly associated with fast developing plant root systems and characteristic for poplar growing under conditions of elevated CO2. Basidomycete - The Basidomycete population importance remained stable; however a decrease in overall population diversity was observed especially among the Homobasidomycetes and included a significant decrease in Heterobasidomycetes, the Tremellomycetidae family most importantly, under elevated CO2. • Goodfellow M. 1983. Ecology of Actinomycetes. Annu Rev Micro 37: 189-216 • Shoun et al. 1998. Denitrification by Actinomycetes and purification of dissimilatory nitrite reductase and azurin from Streptomycesthioluteus. J Bac 180(17): 4413-5 • Sait et al. 2006. Effect of pH on isolation and distribution of members of subdivision 1 of the phylum Acidobacteria occurring in soil. Appl Env Micro 72(3): 1852-7 • Kim et al. 2005. Aerobic nitrification-denitrification by heterotrophic Bacillus strains. Bioresour Technol 96(17): 1897-1906 • Ding et al. 2005. Isolation and identification of nitrogen-fixing bacilli from plant rhizospheres in Beijing region. J Appl Micro 99(5): 1271-81 • Pett-Ridge et al. 2005. Redox fluctuation structures microbial communities in a wet tropical soil. Appl Env Micro 71(11): 6998-7007 • Janssen PH. 2006. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl Env Micro 72(3): 1719-1728 • Minz et al. 1999. Diversity of sunfate reducing bacteriain oxic and anoxic regios of a microbial mat characterized by comparative analysis of dissimilatory sulfite reductase genes. Appl Env Micro 65(10): 4666-4671 • Buckley et al. 2001. Environmental factors influencing the distribution of rRNA from Verrucomicrobia in soil. FEMS Micro Ecol 35(1): 105-112 • Zavaleta et al. 2003. Plants reverse warming effect on ecosystem water balance. PNAS 100(17): 9892-3 • Swann et al. 2001. Urediniomycetes. Pp. 37-56. In: The Mycota VII. Systematics and Evolution. Part B. (Mclaughlin et al. eds.). Springer-Verlag, Berlin. The difference in population importance between the two archaeal populations, as determined by qPCR, was surprising with representatives clustering within this domain decreasing significantly under elevated CO2. The 1500 clones from each library revealed that both populations were predominantly comprised of uncultivated non-thermophillic Crenarchaeota. Members of this domain have previously been documented in soils with some specifically associated with plant roots, suggesting that they may play a significant role in the ecology of the rhizosphere. This work was supported by the US DOE, Office of Science, BER, project entitled New Genomic Strategies and Technologies and under Laboratory Directed Research and Development project number LDRD04-060. We would especially like to thank the JGI for all of their sequencing and Donald Zak at The University of Michigan for providing us with the rhizosphere soil samples. * lesaulnier@bnl.gov