Download

1 / 29
320 likes | 578 Vues
Terminal Oxidative Phosphorylation. NAD + and NADH. Nicotinamide adenine dinucleotide is a central intermediate Previously encountered as a product of glycolysis NADH is formed by the addition of a hydrogen nucleus and 2 electrons (hydride ion) to NAD +

E N D
NAD+ and NADH • Nicotinamide adenine dinucleotide is a central intermediate • Previously encountered as a product of glycolysis • NADH is formed by the addition of a hydrogen nucleus and 2 electrons (hydride ion) to NAD+ • Nicotinamide ring has reduced stability when it accepts the hydride ion since it is no longer stabilized by resonance. • Consequently the electrons (i.e., hydride ion) of NADH can be readily transferred
Example of a reaction involving NAD+ and NADH • Oxidation of an alcohol: two hydrogen atoms are lost. One is added as a hydride ion (H.) to NAD+ the other is released into solution as a proton (H+)
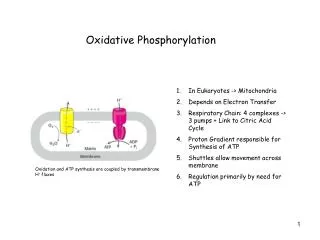
Another view of oxidative phosphorylation • The energetically favorable reaction • H2 + 1/2 O2 ---> H2O • is made to occur in many small steps. • This allows the energy to be stored rather than released as heat. • Electron transport involves a respiratory chain (or electron transport chain). • Respiratory chain consists of protein pumps that harness energy from electron transfer to perform work. • Electrons ultimately are passed to oxygen. • The carriers are transmembrane protein complexes. • The work done is to pump H+ across the inner membrane. This has 2 results: • Low [H+] in the mitochondrial matrix • Voltage gradient - matrix side more negative than outside
Another overview of oxidative phosphorylation • The electrons from H2 (or NADH) are used to reduce O2. • It is evident that the reduction potential of NADH must be higher than the next step in the respiratory chain (or electron transport chain). Only then the acceptor of the electrons can be reduced (accept the electrons). • The same applies to the next steps until the electrons are passed to oxygen which has the lowest reduction potential (but the highest oxidation potential). • In order to better understand the flow of electrons through the respiratory chain it is good to measure the redox potential.
The redox potential (E) • An electrochemical concept that measures the affinity of a substance for electrons • Consider a substance that can exist in an oxidized form X and a reduced form X- (redox couple). • The reduction potential of this couple can be determined by measuring the electromotive force (voltage) generated by a sample half-cell connected to a standard reference half-cell • The sample half-cell consists of an electrode immersed in a solution of 1 M oxidant (X) and 1 M reductant (X-). The standard reference half-cell consists of an electrode immersed in a 1 M H+ solution that is in equilibrium with H2 gas at 1 atmosphere pressure. • The electrodes are connected to a voltmeter, and an agar bridge establishes electrical continuity between the half-cells.
The redox potential (E) • Electrons then flow from one half-cell to the other. If the reaction proceeds in the direction the reactions in the half-cells (referred to as half-reactions or couples) must be • Thus, electrons flow from the sample half-cell to the standard reference half-cell, and the sample-cell electrode is taken to be negative with respect to the standard-cell electrode. The reduction potential of the X:X-couple is the observed voltage at the start of the experiment (when X, X-, and H+ are 1 M). The reduction potential of the H+:H2couple is defined to be 0 volts. • In the example, the reduction potential of the X:X-couple is higher (more negative E) than the reduction potential of the H+:H2couple. The oxidation potential of the X:X-couple is lower (more negative E) than the reduction potential of the H+:H2couple.
Standard reduction potentials of some reactions • If we know the redox potentials of half-reactions, we can predict how a redox reaction will proceed.
Reduction potentials and free energy • The redox potential of reactants/products determines in which direction redox reactions will proceed – but there is also a relation between the difference of the redox potentiaIs of a redox pair and the free energy change: • ΔG°’= -nFΔE°’ • where n is the number of electrons involved and F is the Faraday constant (23 kcal/mol V). • Thus, one can even predict the energy liberated or needed for a reaction. • The redox potential of the NADH/NAD+ pair is –0.32 • The redox potential of the H2O/ ½O2+H+ pair is +0.82 • nFΔE°’ = 1.14 V • ΔG°’ = - (2)(23 kcal/mol V)(1.14 V) • = -52 kcal/mol • ΔG°’ for the formation of ATP is +8 kcal/mol. • Thus, if electron transport chain is 100% efficient it could be predicted that (52/8) or ≈6 ATP would be formed per NADH oxidized. • The highest actual values measured are around 3, or 50% efficiency which is pretty good.
A little history • Once the relationship between the redox potentials of the respiratory chain was worked out, it was easy to identify steps in the process where there was enough energy to phosphorylate ADP, and it was assumed that at these ‘phosphorylation sites’, phosphorylation was coupled directly to the corresponding redox reaction. • So it was possible to calculate a reasonable stoichiometry for the whole process that related Pi/O and thus ATP/O2. The numbers for this were well accepted by 1960. • However, it was seldom possible to obtain the calculated numbers with isolated mitochondria, and never possible to get phosphorylation to happen in fragmented mitochondria or in a soluble system.
Continued… • In 1961 Peter D. Mitchell published a hypothesis for the nature of the coupling between electron transport and phosphorylation – the chemiosmotic hypothesis. • This hypothesis is universally accepted now, and Mitchell received the Nobel Prize for it in 1978. • However, many textbook authors have still not figured out the implications of this mechanism for the yield of ATP from complete oxidation of glucose.
Morals of the story • Much of what you learn in school is not true • Science progresses by revolution rather than steady evolution • Scientists are, on balance, not better about changing their views than non-scientists
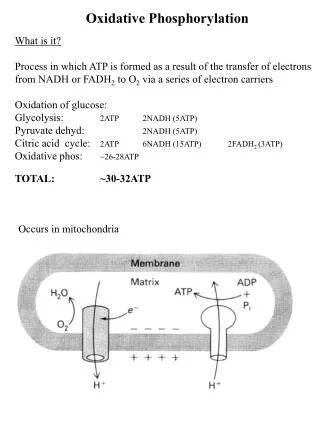
The electron transport chain in the inner mitochondrial membrane • Electron flow through 4 complexes. Some things to take note of: The NADH that feeds into this process can come from glycolysis, lipolysis and/or the TCA cycle. At some points in the sequence that starts with NADH, some of the free energy is used to transport H+ across the inner mitochondrial membrane FADH2 derived from the succinate-fumarate step of the TCA cycle has only enough free energy to energize two of these H+ sites.
Coupling between TCA cycle and oxidative phosphorylation • The reduction equivalents NADH and FADH2 couple the TCA cycle to the oxidative phosphorylation. • NAD+ and FAD+ are reduced in the TCA cycle and their oxidized forms are regenerated in terminal oxidative metabolism, with oxidative phosphorylation of ADP.
A different view • Electron transport energizes H+ transport from the matrix into the intermembrane space. 1 NADH = 10 H+ 1 FADH2 = 6 H+ • FADH2 results in less H+ transport.
Generation of a proton motive force • The osmotic and electrical effects of the proton gradient result in a proton-motive force (PMF or Δp). • pH gradient across the membrane is ΔpH = 1.4 • Voltage across the membrane is 0.14 V (outside positive) • magnitude of Δp can be measured in volts • Δp = 0.14 V - (2.3RT/F)(ΔpH) = 0.224 V • This Δp corresponds to a ΔG of 5.2 kcal per mol of protons
Use of the proton-motive force • ATP generation by ATP synthase or F0F1-ATPase. • F1 unit contains the catalytic site for ATP synthesis • Flow of 3 protons through ATP synthase leads to phosphorylation of 1 ADP • Drive transport of necessary/useful substances.
Transport across inner mitochondrial membrane • ADP and ATP need to be exchanged across the inner mitochondrial membrane • Other transporters:
Cytosolic NADH enters mitochondria via shuttles • Glycerol 3-Phosphate Shuttle This one costs some energy – the difference between an NADH and a FADH2 – and the cytosolic NADH does not literally enter the mitochondria • Malate-Aspartate Shuttle This one operates for free – swapping a NADH in the mito for a NAD+ in the cytoplasm
What is the ATP yield from oxidative phosphorylation? • Yield of ATP (or GTP) from glycolysis or citric acid is unequivocally known • For oxidative phosphorylation, stoichiometry of proton pumping, ATP synthesis, and metabolite transport • need not be integers or have fixed values. • What is the ATP yield based on current estimates? • NADH oxidation ---> 10 H+ pumped • FADH2 oxidation ---> 6 H+ pumped • 3 H+ flow in per 1 ATP produced • ATP/ADP translocase costs 1 H+ per ADP • Therefore 2.5 ATP per NADH, 1.5 ATP per FADH2 • If NADH originated from glycolysis (cytosol), it must be transported into mitochondrial matrix – this may or may not cost something • Pyruvate is transported into the matrix by an OH- coupled transporter, so it is driven by the H+ gradient – this reduces the yield • So ultimately, yield is around 1.5 ATP per cytosolic NADH
What is the actual yield of ATP from oxidative phosphorylation?
Control of terminal oxidative metabolism by ADP • At low activity levels, when ADP levels are low, the proton electrochemical gradient approaches its maximum possible value, so electron flow through the ETC slows, oxygen uptake drops, and levels of reduced coenzyme build up. High ATP levels, low levels of oxidized coenzyme, and low levels of ADP feed back to the substrate-level reactions, slowing them. • If activity levels are increased, ADP arriving at the phosphorylation sites starts the F1 F0 ATPase spinning and as the proton gradient runs down somewhat, it is easier for electrons to flow from reduced coenzymes to oxygen. Levels of reduced coenzymes drop, oxygen consumption rises, and inhibition of the substrate-level reactions is relieved.
Fate of C-skeleton of amino acids • Amino acids that are degraded to acetyl CoA or acetoacetyl CoA are termed ketogenic amino acids (yellow), because they can give rise to ketone bodies or fatty acids. • Amino acids that are degraded to pyruvate, a-ketoglutarate, succinyl CoA, fumarate, or oxaloacetate are termed glucogenic amino acids. The net synthesis of glucose from these amino acids is feasible, because these citric acid cycle intermediates and pyruvate can be converted into phosphoenolpyruvate and then into glucose. • Mammals lack a pathway for the net synthesis of glucose from acetyl CoA or acetoacetyl CoA.
Another thought question • There are animals – such as the housefly – that use oxidative phosphorylation to fuel flight • When a fly takes off, it has to rapidly kick its mitochondria into high gear • Fly blood has high levels of various amino acids, especially proline – how could this relate to the ability to take off quickly?
Oxidative Phosphorylation, Natural Doping, Ecology and Global Warming • The omega-3 forms of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) greatly increase capacity for oxidative metabolism and exercise tolerance. • The mechanism for this involves both a general increase in membrane unsaturated fatty acids and an increase in the expression of enzymes involved in oxidative metabolism and fat metabolism. • The increase in oxidative capacity that results from this diet alone (roughly 20% in a sedentary bird model – the quail) is comparable to what can be achieved with extreme regimes of exercise.
The semipalmated sandpiper migrates from the Canadian and Alaskan Arctic to South America. In preparation for their trip across the Atlantic, the sandpipers stop off in staging areas on the Canadian east coast, where they gorge themselves on a particular species of amphipod, Corophium volutator. In the process, they double their weight in about 2 weeks. The crustaceans they eat contain record amounts of DHA and EPA fatty acids. This dietary strategy apparently makes transoceanic flight possible.
Global warming is causing the natural ranges of the crustaceans and molluscs normally consumed by the birds to shift northward. As a result, the migration on which this species depends may become impossible.