mRNA metabolism
mRNA metabolism. Ülo Maiväli ymaivali@ebc.ee. m 7 G CAP lisatakse kõigile RNA pol II produktidele, osadel protsessitakse hiljem maha. mRNA snRNA snoRNA Telomeraasi RNA. CAPi funktsioonid tuumas (kompleksis CBC-ga). mRNA splaisingu aktiveerimine tuumas

mRNA metabolism
E N D
Presentation Transcript
mRNA metabolism Ülo Maiväli ymaivali@ebc.ee
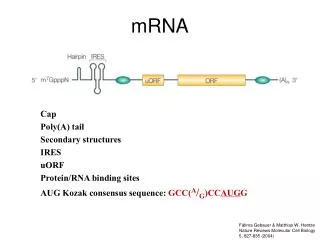
m7G CAP lisatakse kõigile RNA pol II produktidele, osadel protsessitakse hiljem maha • mRNA • snRNA • snoRNA • Telomeraasi RNA
CAPi funktsioonid tuumas (kompleksis CBC-ga) • mRNA splaisingu aktiveerimine tuumas • mRNA 3´-otsa protsessimise/ polüadenülatsiooni aktivatsioon • snRNAde tuuma-tsütoplasma-tuuma transport
snRNAde edasi tagasi liigutamine • m7G CAP eksporditakse tuumast tsütoplasmasse • Seal toimub m7G topeltmetüleerimine 2. positsioonist trimetüülguanosiin süntaas 1 (Tgs1) poolt • m2,2,7G imporditakse tuuma tagasi • m2,2,7G hoiab RNA tuumas
CAPi funktsioonid tsütoplasmas • Toob ribosoomid mRNA-le (üle eIF4F kompleksi). See on translatsiooni kiirust limiteeriv etapp. • Kaitseb 5´-3´ suunalise mRNA eksonukleolüütilise lagundamise eest. PABP interaktsioon CAP-ga selleks vajalik.
mRNA stabiilsus Inimese rakkudes on enamusi mRNAsid u 6 koopiat raku kohta, vaid väheseid 50-100 koopiat. mRNA stabiilsused varieeruvad alates 100X rakutsüklist lühemast poolestusajast kuni mitme rakutsükli pikkuseni. Paljude mRNAde poolestusajad on reguleeritavad. mRNA stabiilsused: varase vastuse geenid (proto-onkogeenid, tsütokiinid) 10-60 min; majahoidjad (histoonid, hormoonid) u 12 h; globiinid, müosiinid, IgG u 24 h. mRNAd siduvad valgud: 1-globaalsed (PABP) 2-grupispetsiifilised (ELAV/Hu) 3-tüübispetsiifilised (IRE). Pärmis 471 annoteeritud RNA-seostuvat valku, millest 312 osalevad RNA protsessingus, modifitseerimistes, splaisingus või lagundamises. Inimeses võib-olla ca 1500 RNA-siduvat valku; palju neist ei tea. inimeses 30 põhilist hnRNP valku ja palju minoorseid.
mRNA lagundamine • 5´ 3´ lagundamine tsütoplasmas • 3´ 5´ lagundamine tsütoplasmas • ARE elemendid- mRNA destabiliseerimine • Lagundamine tuumas • Lagundamine endonukleaasi poolt • NSD- nonstop mediated decay (3´-5´) • NMD- nonsense mediated decay • mRNA lagundamine bakteris
5´ 3´ lagundamine • mRNA deadenüleerimine 10-15 A-ni (Pop2-Ccr4 kompleks, Pan2-Pan3 kompleks, PARN) • CAPi mahavõtmine (Dcp2-Dcp1 kompleks) • 5´-3´ lagundamine exonukleaasi Xrn1 poolt
PolüA inhibeerib mRNA degradatsiooni tänu PABP interaktsioonile. Huvitaval kombel on PABP miinus tüved keskmiselt pikema polüAga. Võib-olla varjab mRNA tsirkulariseerimise kaudu capi ära? Deadenüleeriv ensüüm PARN seondub 5´caple, see stimuleerib aktiivsust. Seega on võimalik et polüA eemaldamise tingimuseks on normaalse mRNA tsirkulariseerimise muutus.
Kui translatsiooni initsatsiooni inhibitsioon destabiliseerib mRNAd, siis elongatsiooni inhibitsioon stabiliseerib seda! PAB1 mutatsiooni viivad prematuursele decappimisele. Mutatsioonid, mis vähendavad decappimist, supresseerivad pab1 deletsiooni letaalsust. Ka eIF4E ja 4G mutatsioonid tõstavad deadenülatsiooni ja decappimist. Dcp1 seob eIF4G-d, siit mudel kus ta vahetab välja eIF4E valgu.
3´ 5´ lagundamine • PolüA lagundamine (PARN) • Tsütoplasmaatiline eksosoom (vähemalt 9 eksonukleaasi) sööb CAPiga mRNA • DcpS hävitab CAP-i järelejäänud lühikese (alla 10 nt) mRNA jupi otsast
Eksosoom koosneb minimaalselt: • 6st RNaas-PH-domääni valgust (PM/SCL75, MTR3, RRP41, RRP42, RRP43 ja RRP46) • 3st S1 ja KH domääni valgust (RRP4, RRP40,CSL4) • RNaasD-sarnasest PM/SCL100 • Helikaas (?) KIAA0053 • Valgust, mida rakutsükli M faasis fosforüülitakse
ARE mRNAd A+U rikas element ARE paljude ebastabiilsete mRNAde 3´-UTRs. 3´-5´ degradatsioonirada põhiline. 8% inimese mRNA-dest sisaldavad ARE elemente. Ei ole selge kuidas ARE-siduvad valgud töötavad. Vist on 1. translatsiooniround vajalik, et ARE mRNAsid destabiliseerida.
ARE-siduvad valgud • AUF1 – destabiliseerib/ kuumaehmatusel stabiliseerib • HuR – stabiliseerib ARE-mRNAd, reguleerib nende tuumaeksporti ja lokalisatsiooni tsütoplasmas • TTP, KSRP, BRF1 - destabiliseerivad
mRNA endonukleolüütiline lagundamine • PMR1- polysomal ribonuclease 1 vahendab östrogeeni poolt stimuleeritud üldist seerumivalkude mRNAde degradatsiooni Xenopuse ootsüüdis. • Tunneb ära AYUGA järjestust. • Osaleb ka NMDs erütrotsüüdis (β-globiini mRNA)
mRNA-d, mille 3´-UTR AU rikas seovad ELAV/Hu valke, mis stabiliseerivad ja/või soodustavad translatsiooni. 3 neljast ELAV/Hu valgust ekspresseeritud neuronis või gonaadides tsütoplasmaatiliselt (dendriitides tsütoplasmaatilisetes graanulites-seal soodustavad translatsiooni: glc-transporteri, neurofilament M translatsioon üles). HuB valk seob vähemalt 100 erinevat mRNAd, peaaegu kõik neist raku kasvu reguleerivate valkude mRNAd, mis sisaldavad AREsid (CREB, c-myc, TNF-alfa jt). Osad AREd võivad vahest hoopis mRNAd stabiliseerida.
mRNA lagundamine tuumas • Tuuma eksosoom 3´-5´- unikaalne komponent Rrp6p (pärm). Vajalik snRNA, rRNA, snoRNA, pre-mRNA maturatsiooniks ja RNA degradatsiooniks tuumas. • Rrp6p osaline kvaliteedikontrollil värskelt sünteesitud mRNA vabastamisel transkriptsioonikohast. • Ka tuumaeksosoom võib ARE-mRNAd lagundada
NSD- nonstop mediated decay • Polü(A) translatsioon lisab valgule polü(Lys) saba • Ribosoom jääb otsa kinni, seob Ski7 (eRF3 homoloog) • Ski7 stimuleerib ribosoomi dissotsatsiooni ja toob eksosoomi kohale
NMD- nonsense mediated decaytoimub kõigis eukarüootides • Enneaegsete stop-koodonitega mRNAd • Upstream ORF in 5´UTR • Liiga pikk 3´UTR • Bitsistroonsed mRNAd • mRNAd kus kasutatakse korraga mitmeid eri raamides start koodoneid
NMD on tähtis protsess • 10% pärmi mRNAde tase muutub kui NMD-d inhibeerida • 1/3 imetaja alternatiivsest splaisingust toodab NMD substraate • NMDs inaktiivsed hiire embrüod surevad varsti pärast implantatsiooni • NMDs inaktiivsed blastotsüstid lähevad kultuuris apoptoosi
Miks NMD?Pooliku ORFiga mRNAlt peaks transleeritama poolikut valku. Poolikud valgud, mis näiteks on jäänud ilma olulistest domäänidest võivad kahjustada organismi elutegevust töötades näiteks onkogeenidena või viies dominantselt negatiivsele fenotüübile. On teada üle 200 enneaegset stop-koodonit tekitava mutatsiooni, mis põhjustavad haigusi (enneaegse stop-koodoni tekkele viib ka enamus raaminihet põhjustavaid mutasioone). Kui NMD-d ei oleks, oleksid paljud retsessiivsed mutatsioonid dominantse mõjuga (ja põhjustaksidraskemaid haigusi).
NMD kohta 2 olulist fakti: 1) NMD toimub enamasti enne kui on katkenud mRNA füüsiline side tuumaga. Enneaegset stop-koodonit sisaldav mRNA molekul, mis juba ilusti tsütoplasmas ujub, pääseb suure tõenäosusega lagundamisest.
2) NMD on alati seotud lagundatava mRNA translatsiooniga. Enneaegset stop-koodonit sisaldava mRNA translatsiooni inhibeerimine stabiliseerib vastavat mRNAd. Konkreetselt on NMD toimumiseks vajalik translatsiooni terminatsioon enneaegsel stop-koodonil (supressor-tRNAd inhibeerivad NMD-d).
Translatsioon, mille toimumisest NMD sõltub, initseeritakse sõltuvalt tuumas cap-i siduvatest valkudest CBP80 ja CBP20. Kuna juba peale esimest translatsioonitsüklit vahetatakse need valgud mRNA 5´-cap struktuuril eIF4E vastu, toimub NMD peale esimest ja enne teist translatsioonitsüklit.
Imetajas on NMD toimumiseks vaja, et stop koodononile kus translatsioon termineeritakse järgneks rohkem kui 50-55 nt allavoolu intron.
Kuid intron splaissitakse välja tuumas enne seda kui ribosoom saab tsütoplasmas sotp koodonil termineerida!
Tuumas toimuv splaissing jätab 20-24 nt splaisingu toimumiskohast ülesvoolu maha valgulise kompleksi- EJC (exon-junction complex)
ECJ transporditakse tsütoplasmasse, seal eemaldatakse ta mRNAlt esimese translatsioonitsükli käigus.
Esimene translatsioonitsükkel (võimalik, et vaid ühe ribosoomi poolt läbi viidud ühe valgumolekuli süntees) toimub arvatavasti veel siis kui mRNA on veel tuumamembraaniga seotud ja on kindlasti seotud CBP80-CBP20 CAPi siduva heterodimeeriga.
Upf valgud on vajalikud NMD-ks • Upf3 seob EJC-le tuumas • Upf2 seob Upf3-le transpordil tsütoplasmasse • Kui transleeriv ribosoom EJC-d ära ei korista, toob eRF3 endaga seotud Upf1 kohale • mRNA lagundatakse.
Upf 1 fosforüülimine • Tavaliselt rakus Upf1 hüpofosforüülitud • Polüsoomides hüperfosforüülitud • Upf1 fosforüülimine (SMG1) ja defosforüülimine (SMG6) NMD-ks vajalik
EJC ja translatsioon • Splaissitud mRNA pealt sünteesitakse rohkem valku kui identse järjestusega splaissimata (cDNAlt transkribeeritud) mRNA pealt. • EJC valgud (RNPS1, Y14, Magoh) suurendavad translatsiooni saagist kui nad kunstlikult mRNAle siduda. • Splaissitud mRNA on palju enam polüsoomidega seotud kui identne splaissimata mRNA. • Süntees splaissitud mRNAlt võib ka valgu stabiilsust tõsta!!! (dihydrolate reductase -2,7x)
Pärmis introneid vähe, seal pole ka EJC kompleksi põhikomponente (Y14, magoh, eIF4AIII). Aga NMD toimub.
Pärmis DSE- downstream sequence element, mis täidab EJC funktsiooni kui asub stop koodonist 3´-suunas.
Drosophilas asi segane: kuigi kõik EJC komponendid olemas, pole ükski neist NMD-ks vajalik.
C. elegansis mängivad Upf1, SMG5 ja SMG6 ka RNA interferentsis mingit (tundmatut) funktsionaalset rolli
mRNA lagundamine bakteris • Keskmise mRNA (5 kb) transktiptsioon 2 min • Translatsioon samuti 2 min (180 kDa valk) • Transkriptsiooni indutseerimisest mRNA tekkeni u 2,5 min • Transkriptsiooni indutseerimisest valgu tekkeni u 3 min • Translatsioon efektiivne- tryptofaani operoni transkribeeritakse 15/min ja 30 ribosoomi/mRNA
Lagundamine algab 1min peale sünteesi algust 5´-otsast. Seega enne kui 3´-ots valmis on! • E. colis üle 20 RNaasi, millest 6 osalevad mRNA degradatsioonil • Degradatsioon 5´-3´ suunaline, u 2X aeglasem kui translatsioon • Bakteris ei ole 5´-3´ eksonukleaasset aktiivsust. • Seal endonukleaasid (initseerivad degradatsiooni lõigates viimasest ribosoomist 5´-suunas: seega lagundamise üldine suund ikkagi 5´-3´). • Seejärel 3´-5´ eksonukleaasid lagundavad endo poolt genereeritud jupid
Keskmine mRNA poolestusaeg 2 min (30´´-20´) • Endonukleaasile tundlikud kohad destabiliseerivad mRNAd • mRNA struktuurid, mis pidurdavad eksonukleaase stabiliseerivad
Degradosoom:1.5-2.4x103 kDa Põhikomponendid: • RNase E – 5´-otsast sõltuv ssRNA-spetsiifiline endonukleaas (ds ots või trifosfaat inhibeerib) • PNPase – 3´-5´ eksonukleaas • RhlB – DEAD box RNA helikaas Lisaks substöhhiomeetriliselt: • Enolaas, polüfosfaadi kinaas, DnaK, GroEL, polü(A) polümeraas, r-valk S1
Minimaalne degradosoom • RNase E dimeer • PNPase trimeer • RhlB dimeer
Degradosoomi funktsioonid: • mRNA lagundamine • 5S rRNA protsessimine • rRNA lagundamine